Wolbachia
| Wolbachia | |
|---|---|
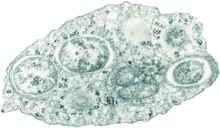 | |
| Transmission electron micrograph of Wolbachia within an insect cell. Credit:Public Library of Science / Scott O'Neill | |
| Scientific classification | |
| Domain: | Bacteria |
| Phylum: | Proteobacteria |
| Class: | Alphaproteobacteria |
| Subclass: | Rickettsidae |
| Order: | Rickettsiales |
| Family: | Rickettsiaceae |
| Genus: | Wolbachia |
| Species[1] | |
|
Wolbachia melophagi (Nöller 1917) Philip 1956[2] Wolbachia persica Suitor and Weiss 1961[3] Wolbachia pipientis Hertig 1936 | |
Wolbachia is a genus of bacteria which infects arthropod species, including a high proportion of insects, as well as some nematodes. It is one of the world's most common parasitic microbes and is possibly the most common reproductive parasite in the biosphere. Its interactions with its hosts are often complex, and in some cases have evolved to be mutualistic rather than parasitic. Some host species cannot reproduce, or even survive, without Wolbachia infection. One study concluded that more than 16% of neotropical insect species carry bacteria of this genus,[4] and as many as 25 to 70 percent of all insect species are estimated to be potential hosts.[5]
History
The genus was first identified in 1924 by Marshall Hertig and S. Burt Wolbach in Culex pipiens, the common house mosquito. Hertig formally described the species in 1936 as Wolbachia pipientis.[6] Research on Wolbachia intensified after 1971, when Janice Yen and A. Ralph Barr of UCLA discovered that Culex mosquito eggs were killed by a cytoplasmic incompatibility when the sperm of Wolbachia-infected males fertilized infection-free eggs.[7][8] The genus Wolbachia is of considerable interest today due to its ubiquitous distribution, its many different evolutionary interactions, and its potential use as a biocontrol agent.
Method of sexual differentiation in hosts
These bacteria can infect many different types of organs, but are most notable for the infections of the testes and ovaries of their hosts. Wolbachia species are ubiquitous in mature eggs, but not mature sperm. Only infected females therefore pass the infection on to their offspring. Wolbachia maximize their spread by significantly altering the reproductive capabilities of its hosts, with four different phenotypes:
- Male killing: infected males die during larval development, which increase the rate of born, infected, females.[9]
- Feminization: infected males develop as females or infertile pseudo-females.
- Parthenogenesis: reproduction of infected females without males. Some scientists have suggested that parthenogenesis may always be attributable to the effects of Wolbachia.[10] An example of a parthenogenic species is the Trichogramma wasp,[11] which has evolved to procreate without males with the help of Wolbachia. Males are rare in this tiny species of insect, possibly because many have been killed by that very same strain of Wolbachia.[12]
- Cytoplasmic incompatibility: the inability of Wolbachia-infected males to successfully reproduce with uninfected females or females infected with another Wolbachia strain. In the mechanism of cytoplasmic incompatibility, Wolbachia interferes with the parental chromosomes during the first mitotic divisions to the extent that they can no longer divide in sync.[13]
Effects of sexual differentiation in hosts
Several species, such as Trichogramma, are so dependent on sexual differentiation of Wolbachia that they are unable to reproduce effectively without the bacteria in their bodies, and some might even be unable to survive uninfected.[14]
One study on infected woodlice showed the broods of infected organisms had a higher proportion of females than their uninfected counterparts.[15]
Wolbachia, especially Wolbachia-caused cytoplasmic incompatibility, may be important in promoting speciation.[16][17][18] Wolbachia strains that distort the sex ratio may alter their host's pattern of sexual selection in nature,[19][20] and also engender strong selection to prevent their action, leading to some of the fastest examples of natural selection in natural populations.[21]
The male killing and feminization effects of Wolbachia infections can also lead to speciation in their hosts. For example, populations of the pill woodlouse, Armadillidium vulgare, which are exposed to the feminizing effects of Wolbachia have been known to lose their female-determining chromosome.[22] In these cases, only the presence of Wolbachia can cause an individual to develop into a female.[22]
Fitness advantages by Wolbachia infections
Wolbachia has been linked to viral resistance in Drosophila melanogaster, Drosophila simulans, and mosquito species. Flies infected with the bacteria are more resistant to RNA viruses such as Drosophila C virus, Nora virus, Flock house virus, Cricket paralysis virus, Chikungunya virus, and West Nile virus.[23][24][25]
In the common house mosquito, higher levels of Wolbachia density were correlated with more insecticide resistance.[26]
In leafminers of the species Phyllonorycter blancardella, Wolbachia bacteria help their hosts produce green islands on yellowing tree leaves, that is, small areas of leaf remaining fresh, allowing the hosts to continue feeding while growing to their adult forms. Larvae treated with tetracycline, which kills Wolbachia, lose this ability and subsequently only 13% emerge successfully as adult moths.[27]
The Muscidifurax uniraptor parasitoid wasp also benefits from hosting a Wolbachia bacteria. Faculty from the Hebrew University of Jerusalem studied the positive effects of Wolbachia on the fecundity of a strain of M. uniraptor from Rochester, New York, in 1997.[28]
In the parasitic filarial nematode species responsible for elephantiasis, such as Brugia malayi and Wuchereria bancrofti, Wolbachia has become an obligate endosymbiont and provides the host with chemicals necessary to its reproduction and survival.[29] Elimination of the Wolbachia symbionts through antibiotic treatment therefore prevent reproduction of the nematode, and eventually results in its premature death.
Some Wolbachia that infect arthropods also provide some metabolic provisioning to their host. In Drosophila melanogaster, Wolbachia is found to mediate iron metabolism under nutritional stress[30] and in Cimex lectularis, Wolbachia strain cCle helps the host to synthesize B vitamins.[31]
Wolbachia is also found to provide the host with a benefit of increasing fecundity. Weeks et al. found that the Wolbachia strains captured from 1988 in southern California still induce a fecundity deficit, but nowadays the fecundity deficit is replaced with a fecundity advantage such that infected Drosophila simulans produces more offspring than the uninfected ones.[32]
Genomics
The first Wolbachia genome to be determined was that of one that infects Drosophila melanogaster flies.[33] This genome was sequenced at The Institute for Genomic Research in a collaboration between Jonathan Eisen and Scott O'Neill. The second Wolbachia genome to be determined was one that infects Brugia malayi nematodes.[34] Genome sequencing projects for several other Wolbachia strains are in progress. A nearly complete copy of the Wolbachia genome sequence was found within the genome sequence of the fruit fly Drosophila ananassae and large segments were found in 7 other Drosophila species.[35]
In an application of DNA barcoding to the identification of species of Protocalliphora flies, it was found that several distinct morphospecies had identical cytochrome c oxidase I gene sequences, most likely through horizontal gene transfer by Wolbachia species as they jump across host species.[36] As a result, Wolbachia can cause misleading results in molecular cladistical analyses.[37]
Small RNA
Wolbachia's small non-coding RNAs WsnRNA-46 and WsnRNA-59 were detected in Aedes aegypti mosquitoes and Drosophila melanogaster. The small RNAs may regulate bacterial and host genes.[38]
Horizontal gene transfer
Wolbachia also harbor a temperate bacteriophage called WO.[39] Comparative sequence analyses of bacteriophage WO offer some of the most compelling examples of large-scale horizontal gene transfer between Wolbachia coinfections in the same host.[40] It is the first bacteriophage implicated in frequent lateral transfer between the genomes of bacterial endosymbionts. Gene transfer by bacteriophages could drive significant evolutionary change in the genomes of intracellular bacteria that were previously considered highly stable or prone to loss of genes overtime.[40]
Applications to human-related infections
Outside of insects, Wolbachia infects a variety of isopod species, spiders, mites, and many species of filarial nematodes (a type of parasitic worm), including those causing onchocerciasis ("River Blindness") and elephantiasis in humans as well as heartworms in dogs. Not only are these disease-causing filarial worms infected with Wolbachia, but Wolbachia seem to play an inordinate role in these diseases. A large part of the pathogenicity of filarial nematodes is due to host immune response toward their Wolbachia. Elimination of Wolbachia from filarial nematodes generally results in either death or sterility of the nematode.[41] Consequently, current strategies for control of filarial nematode diseases include elimination of their symbiotic Wolbachia via the simple doxycycline antibiotic, rather than directly killing the nematode with far more toxic anti-nematode medications.[42]
Naturally existing strains of Wolbachia have been shown to be a route for vector control strategies because of their presence in arthropod populations, such as mosquito populations.[43][44] Due to the unique traits of Wolbachia that cause cytoplasmic incompatibility, this strain is useful as a promoter of genetic drive within a population. Wolbachia-infected females are able to produce offspring with non-infected and infected males, however, non-infected females are only able to produce viable offspring with non-infected males. This gives infected females a frequency-dependent reproductive advantage; that is, the advantage is greater the higher the frequency of Wolbachia in the population. Computational models predict that introducing Wolbachia strains into natural populations will reduce pathogen transmission and reduce overall disease burden.[45] An example includes Wolbachia that can be used to control dengue and malaria by eliminating the older insects that contain more parasites. Promoting the survival and reproduction of younger insects lessens selection pressure for evolution of resistance.[46][47] Wolbachia strains that are able to reduce dengue transmission include wAllbB and wMelPop with Aedes aegypti, wMel with Aedes albopictus.[48] and Aedes aegypti.[49] In addition to inhibit the DENV (Dengue Virus) in some species of genus Aedes, Wolbachia has been identified to inhibit replication of CHIKV (Chikungunya virus) in Aedes aegypti. It was found that Wmel strain of Wolbachia pipientis reduced significantly infection and dissemination rates of CHIKV in mosquitoes, compared to Wolbachia uninfected controls and the same phenomenon was observed in YFV (Yellow Fever Virus) infection converting this bacteria in an excellent promise for YFV and CHIKV suppression.[50]
Another study tested the effect of Wolbachia on the replication of WNV (West Nile Virus) but in cell line Aag2 derived from Aedes aegypti cells, suggesting that despite the existence of an enhancement of viral genomic RNA replication in the cell line Wolbachia infected, the production of secreted virus was significantly inhibited. It also was found that the antiviral effect in intrathoracically infected mosquitoes depends on the strain of Wolbachia, and the replication of the virus in orally fed mosquitoes was completely inhibited in wMelPop strain of Wolbachia.[51]
However, cases where Wolbachia reduces transmission of pathogens must be studied carefully. Indeed, a recent study demonstrated that the West Nile virus could be improved by the presence of the Wolbachia strain wAlbB in the mosquito Culex tarsalis. The rate of virus appears significantly higher than in uninfected mosquitoes. Indeed, wAlbB inhibits REL1, which is an activator of the antiviral Toll immune pathway. The release of artificially infected mosquitoes in the environment, for a vector-borne disease control program, should be made with caution, because it is the first case of improvement of a human disease in the mosquito.[52]
It has been suggested that Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll (gene family) pathway. This pathway is essential for activation of antimicrobial peptides, defensins, and cecropins that help to inhibit dengue virus proliferation.[53] Wolbachia infection can also increase mosquito resistance to malaria, as shown in Anopheles stephensi where the wAlbB strain of Wolbachia hindered the life cycle of Plasmodium falciparum.[54]
Some experts have proposed to combat the spread of the Zika virus by breeding and releasing mosquitoes that have intentionally been infected with an appropriate strain of Wolbachia.[55] A recent study has shown that Wolbachia has the ability to block the spread of Zika virus in mosquitos in Brazil [56]
In October 2016, it was announced that $18 million in funding was being allocated for the use of Wolbachia-infected mosquitoes to fight Zika and dengue virus. Deployment is slated for early 2017 in Colombia and Brazil.[57]
See also
References
- ↑ Most Wolbachia species cannot be cultured outside of their eukaryotic host and so have not been given formal latin names.
- ↑ Arguing that W. melophagi should be transferred to the genus Bartonella
- Dumler JS, Barbet AF, Bekker CP, et al. (November 2001). "Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and 'HGE agent' as subjective synonyms of Ehrlichia phagocytophila". Int. J. Syst. Evol. Microbiol. 51 (Pt 6): 2145–65. doi:10.1099/00207713-51-6-2145. PMID 11760958.
- Lo N, Paraskevopoulos C, Bourtzis K, et al. (March 2007). "Taxonomic status of the intracellular bacterium Wolbachia pipientis". Int. J. Syst. Evol. Microbiol. 57 (Pt 3): 654–7. doi:10.1099/ijs.0.64515-0. PMID 17329802.
- ↑ Arguing that W. persica should be transferred to the genus Francisella
- Forsman M, Sandström G, Sjöstedt A (January 1994). "Analysis of 16S ribosomal DNA sequences of Francisella strains and utilization for determination of the phylogeny of the genus and for identification of strains by PCR". Int. J. Syst. Bacteriol. 44 (1): 38–46. doi:10.1099/00207713-44-1-38. PMID 8123561.
- Noda H, Munderloh UG, Kurtti TJ (October 1997). "Endosymbionts of ticks and their relationship to Wolbachia spp. and tick-borne pathogens of humans and animals". Appl. Environ. Microbiol. 63 (10): 3926–32. PMC 168704
 . PMID 9327557.
. PMID 9327557. - Niebylski ML, Peacock MG, Fischer ER, Porcella SF, Schwan TG (October 1997). "Characterization of an endosymbiont infecting wood ticks, Dermacentor andersoni, as a member of the genus Francisella". Appl. Environ. Microbiol. 63 (10): 3933–40. PMC 168705. PMID 9327558.
- ↑ Werren, J.H.; Guo, L; Windsor, D. W. (1995). "Distribution of Wolbachia among neotropical arthropods". Proceedings of the Royal Society B. 262 (1364): 197–204. Bibcode:1995RSPSB.262..197W. doi:10.1098/rspb.1995.0196.
- ↑ Kozek, Wieslaw J.; Rao, Ramakrishna U. (2007). "The Discovery of Wolbachia in Arthropods and Nematodes – A Historical Perspective". Issues in Infectious Diseases. Issues in Infectious Diseases. 5 (Wolbachia: A Bug’s Life in another Bug): 1–14. doi:10.1159/000104228. ISBN 3-8055-8180-7.
- ↑ Hertig, Marshall; Wolbach, S. Burt (1924). "Studies on Rickettsia-Like Micro-Organisms in Insects". Journal of Medical Research. 44 (3): 329–74. PMC 2041761. PMID 19972605.
- ↑ Yen JH; Barr AR (1971). "New hypothesis of the cause of cytoplasmic incompatibility in Culex pipiens". Nature. 232 (5313): 657–8. Bibcode:1971Natur.232..657Y. doi:10.1038/232657a0. PMID 4937405.
- ↑ Bourtzis, Kostas; Miller, Thomas A. (eds.). "14: Insect pest control using Wolbachia and/or radiation". Insect Symbiosis. p. 230. ISBN 9780849341946.
- ↑ Hurst G., Jiggins F. M., Graf von Der Schulenburg J. H., Bertrand D., et al. (1999). "Male killing Wolbachia in two species of insects". Proceedings of the Royal Society B. 266 (1420): 735–740. doi:10.1098/rspb.1999.0698.
- ↑ Tortora, Gerard J.; Funke, Berdell R.; Case, Cristine L. (2007). Microbiology: an introduction. Pearson Benjamin Cummings. ISBN 0-8053-4790-9.
- ↑ Knight J (5 July 2001). "Meet the Herod Bug". Nature. 421 (6842): 12–14. doi:10.1038/35083744. PMID 11452274.
- ↑ Murray, Todd. "Garden Friends & Foes: Trichogramma Wasps". Weeder's Digest. Washington State University Whatcom County Extension. Retrieved 16 July 2009.
- ↑ Breeuwer, JAJ; Werren, JH (1990). "Microorganisms associated with chromosome destruction and reproductive isolation between two insect species" (PDF). Nature. 346: 558–560. doi:10.1038/346558a0. PMID 2377229.
- ↑ Werren, John H. (February 2003). "Invasion of the Gender Benders: by manipulating sex and reproduction in their hosts, many parasites improve their own odds of survival and may shape the evolution of sex itself" (Reprint). Natural History. 112 (1): 58. ISSN 0028-0712. OCLC 1759475. Retrieved 15 November 2008.
- ↑ Rigaud, Thierry; Moreau, Jérôme; Juchault, Pierre (October 1999). "Wolbachia infection in the terrestrial isopod Oniscus asellus: sex ratio distortion and effect on fecundity". Heredity. 83 (4): 469–475. doi:10.1038/sj.hdy.6885990. PMID 10583549. However, the broods also often consisted of fewer eggs than the broods of the uninfected Oniscus asellus.
- ↑ Bordenstein, Seth R.; Patrick, O'Hara; Werren, John H. (2001). "Wolbachia-induced incompatibility precedes other hybrid incompatibilities in Nasonia". Nature. 409 (6821): 675–7. Bibcode:2001Natur.409..707B. doi:10.1038/35055543. PMID 11217858.
- ↑ Zimmer, Carl (2001). "Wolbachia: A Tale of Sex and Survival". Science. 292 (5519): 1093–5. doi:10.1126/science.292.5519.1093. PMID 11352061.
- ↑ Telschow, Arndt; Flor, Matthias; Kobayashi, Yutaka; Hammerstein, Peter; Werren, John H. (2007). Rees, Mark, ed. "Wolbachia-Induced Unidirectional Cytoplasmic Incompatibility and Speciation: Mainland-Island Model". PLoS ONE. 2 (1): e701. Bibcode:2007PLoSO...2..701T. doi:10.1371/journal.pone.0000701. PMC 1934337. PMID 17684548.
- ↑ Charlat S., Reuter M., Dyson E.A., Hornett E.A., Duplouy A.M.R., Davies N., Roderick G., Wedell N., Hurst G.D.D. (2006). "Male-killing bacteria trigger a cycle of increasing male fatigue and female promiscuity". Current Biology. 17 (3): 273–7. doi:10.1016/j.cub.2006.11.068. PMID 17276921.
- ↑ JIGGINS F. M.; Hurst, G. D. D.; Majerus, M. E. N. (2000). "Sex ratio distorting Wolbachia cause sex role reversal in their butterfly hosts". Proceedings of the Royal Society B. 267 (1438): 69–73. doi:10.1098/rspb.2000.0968.
- ↑ Charlat, S.; Hornett, E. A.; Fullard, J. H.; Davies, N.; Roderick, G. K.; Wedell, N.; Hurst, G. D. D. (2007). "Extraordinary Flux in Sex Ratio". Science. 317 (5835): 214. Bibcode:2007Sci...317..214C. doi:10.1126/science.1143369. PMID 17626876.
- 1 2 Charlat, Sylvain (April 2003). "Evolutionary consequences of Wolbachia infections" (PDF). Trends in Genetics. 19: 217–23. doi:10.1016/S0168-9525(03)00024-6. PMID 12683975. Retrieved September 28, 2015.
- ↑ Teixeira, Luis; Ferreira, Alvaro; Ashburner, Michael (2008). Keller, Laurent, ed. "The Bacterial Symbiont Wolbachia Induces Resistance to RNA Viral Infections in Drosophila melanogaster". PLOS Biology. 6 (12): e2. doi:10.1371/journal.pbio.1000002. PMC 2605931. PMID 19222304.
- ↑ Hedges, Lauren; Brownlie, Jeremy; O'Neill, Scott; Johnson, Karyn (2008). "Wolbachia and Virus Protection in Insects". Science. 322 (5902): 702. Bibcode:2008Sci...322..702H. doi:10.1126/science.1162418.
- ↑ Glaser, Robert L.; Meola, Mark A. (2010). Liu, Ding Xiang, ed. "The Native Wolbachia Endosymbionts of Drosophila melanogaster and Culex quinquefasciatus Increase Host Resistance to West Nile Virus Infection". PLOS Biology. 5 (6): e11977. Bibcode:2010PLoSO...511977G. doi:10.1371/journal.pone.0011977. PMC 2916829. PMID 20700535.
- ↑ Berticat C, Rousset F, Raymond M, Berthomieu A, Weill M (July 2002). "High Wolbachia density in insecticide-resistant mosquitoes". Proc. Biol. Sci. 269 (1498): 1413–6. doi:10.1098/rspb.2002.2022. PMC 1691032. PMID 12079666.
- ↑ Kaiser W, Huguet E, Casas J, Commin C, Giron D (August 2010). "Plant green-island phenotype induced by leaf-miners is mediated by bacterial symbionts". Proc. Biol. Sci. 277 (1692): 2311–9. doi:10.1098/rspb.2010.0214. PMC 2894905. PMID 20356892.
- ↑ Zchori-Fein, Einat (2000). "Wolbachia Density and Host Fitness Components in Muscidifurax uniraptor (Hymenoptera: Pteromalidae)" (PDF). Journal of Invertebrate Pathology. 75: 267–272. doi:10.1006/jipa.2000.4927. Retrieved September 22, 2015.
- ↑ Foster, Jeremy; Ganatra, Mehul; Kamal; Ware, Jennifer; Makarova, Kira; Ivanova, Natalia; Bhattacharyya, Anamitra; Kapatral, Vinayak; et al. (2005). "The Wolbachia Genome of Brugia malayi: Endosymbiont Evolution within a Human Pathogenic Nematode". PLOS Biology. 3 (4): e121. doi:10.1371/journal.pbio.0030121. PMC 1069646. PMID 15780005.
- ↑ Brownlie, J.C.; Cass, B. N.; Riegler, M. (2009). "Evidence for metabolic provisioning by a common invertebrate endosymbiont, Wolbachia pipientis, during periods of nutritional stress". PLOS Pathogens. 5 (4): e1000368. doi:10.1371/journal.ppat.1000368.
- ↑ Nikoh, N.; Hosokawa, T.; Moriyama, M. (2014). "Evolutionary origin of insect- Wolbachia nutritional mutualism". Proceedings of the National Academy of Sciences of the United States of America. 111 (28): 10257–10262. doi:10.1073/pnas.1409284111.
- ↑ Weeks, A.R.; Turelli, M.; Harcombe, W.R. (2007). "From parasite to mutualist: rapid evolution of Wolbachia in natural populations of Drosophila". PLOS Biology. 5 (5): e114. doi:10.1371/journal.pbio.0050114.
- ↑ Wu M, Sun LV, Vamathevan J, et al. (2004). "Phylogenomics of the Reproductive Parasite Wolbachia pipientis wMel: A Streamlined Genome Overrun by Mobile Genetic Elements". PLOS Biology. 2 (3): e69. doi:10.1371/journal.pbio.0020069. PMC 368164. PMID 15024419.
- ↑ Foster J, Ganatra M, Kamal I, et al. (2005). "The Wolbachia Genome of Brugia malayi: Endosymbiont Evolution within a Human Pathogenic Nematode". PLOS Biology. 3 (4): e121. doi:10.1371/journal.pbio.0030121. PMC 1069646. PMID 15780005.
- ↑ Dunning Hotopp, J.C, Clark ME, Oliveira DC, Foster JM, Fischer P, Torres MC, Giebel JD, Kumar N, Ishmael N, Wang S, Ingram J, Nene RV, Shepard J, Tomkins J, Richards S, Spiro DJ, Ghedin E, Slatko BE, Tettelin H, Werren J.H.; Clark; Oliveira; Foster; Fischer; Torres; Giebel; Kumar; Ishmael; Wang; Ingram; Nene; Shepard; Tomkins; Richards; Spiro; Ghedin; Slatko; Tettelin; Werren (2007). "Widespread Lateral Gene Transfer from Intracellular Bacteria to Multicellular Eukaryotes". Science. 317 (5845): 1753–6. Bibcode:2007Sci...317.1753H. doi:10.1126/science.1142490. PMID 17761848.
- ↑ Whitworth, TL; Dawson, RD; Magalon, H; Baudry, E (2007). "DNA barcoding cannot reliably identify species of the blowfly genus Protocalliphora (Diptera: Calliphoridae)". Proceedings of the Royal Society B. 274 (1619): 1731–9. doi:10.1098/rspb.2007.0062. PMC 2493573. PMID 17472911.
- ↑ Johnstone, RA; Hurst, GDD (1996). "Maternally inherited male-killing microorganisms may confound interpretation of mitochondrial DNA variability". Biological Journal of the Linnean Society. 58 (4): 453–470. doi:10.1111/j.1095-8312.1996.tb01446.x.
- ↑ Mayoral, Jaime G.; Hussain, Mazhar; Joubert, D. Albert; Iturbe-Ormaetxe, Iñaki; O'Neill, Scott L.; Asgari, Sassan (2014-12-30). "Wolbachia small noncoding RNAs and their role in cross-kingdom communications". Proceedings of the National Academy of Sciences of the United States of America. 111 (52): 18721–18726. doi:10.1073/pnas.1420131112. ISSN 1091-6490. PMC 4284532. PMID 25512495.
- ↑ Masui S, Kamoda S, Sasaki T, Ishikawa H (2000). "Distribution and evolution of bacteriophage WO in Wolbachia, the endosymbiont causing sexual alterations in arthropods.". J Mol Evol. 51 (5): 491–491. doi:10.1007/s002390010112. PMID 11080372.
- 1 2 Kent, Bethany N.; Salichos, Leonidas; Gibbons, John G.; Rokas, Antonis; Newton, Irene L.; Clark, Michael E.; Bordenstein, Seth R. (2011). "Complete Bacteriophage Transfer in a Bacterial Endosymbiont (Wolbachia) Determined by Targeted Genome Capture". Genome in Biology and Evolution. 3: 209–218. doi:10.1093/gbe/evr007. PMC 3068000. PMID 21292630.
- ↑ Hoerauf A, Mand S, Fischer K, et al. (2003). "Doxycycline as a novel strategy against bancroftian filariasis-depletion of Wolbachia endosymbionts from Wuchereria bancrofti and stop of microfilaria production". Med. Microbiol. Immunol. 192 (4): 211–6. doi:10.1007/s00430-002-0174-6. PMID 12684759.
- ↑ Taylor, MJ; Makunde, WH; McGarry, HF; Turner, JD; Mand, S; Hoerauf, A (2005). "Macrofilaricidal activity after doxycycline treatment of Wuchereria bancrofti: a double-blind, randomised placebo-controlled trial". Lancet. 365 (9477): 2116–21. doi:10.1016/S0140-6736(05)66591-9. PMID 15964448.
- ↑ Xi, Z; Dean JL; Khoo C; Dobson SL. (2005). "Generation of a novel Wolbachia infection in Aedes albopictus (Asian tiger mosquito) via embryonic microinjection". Insect Biochem Mol Biol. 35 (8): 903–10. doi:10.1016/j.ibmb.2005.03.015. PMC 1410910. PMID 15944085.
- ↑ Moreira, LA; Iturbe-ormaetxe I; Jeffery JA; et al. (2009). "A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium". Cell. 139 (7): 1268–78. doi:10.1016/j.cell.2009.11.042. PMID 20064373.
- ↑ Hancock, PA; Sinkins SP; Godfray HC. (2011). "Strategies for introducing Wolbachia to reduce transmission of mosquito-borne diseases". PLOS Negl. Trop. Dis. 5 (4): e1024. doi:10.1371/journal.pntd.0001024. PMC 3082501. PMID 21541357.
- ↑ Mcmeniman, CJ; Lane RV; Cass BN; et al. (2009). "Stable introduction of a life-shortening Wolbachia infection into the mosquito Aedes aegypti". Science. 323 (5910): 141–4. doi:10.1126/science.1165326. PMID 19119237.
- ↑ "'Bug' could combat dengue fever". BBC NEWS. British Broadcasting Corporation. 2 January 2009.
- ↑ Blagrove, MS; Arias-goeta C; Failloux AB; Sinkins SP. (2012). "Wolbachia strain wMel induces cytoplasmic incompatibility and blocks dengue transmission in Aedes albopictus". Proc Natl Acad Sci U S A. 109 (1): 255–60. doi:10.1073/pnas.1112021108. PMC 3252941. PMID 22123944.
- ↑ Hoffmann, AA; Iturbe-Ormaetxe I; Callahan AG; Phillips BL. (2014). "Stability of the wMel Wolbachia Infection following invasion into Aedes aegypti populations". PLOS Negl. Trop. Dis. 8: e3115. doi:10.1371/journal.pntd.0003115.
- ↑ van den Hurk, AF; Hall-Mendelin S; Pyke AT; Frentiu FD. (2012). "Impact of Wolbachia on infection with chikungunya and yellow fever viruses in the mosquito vector Aedes aegypti". PLOS Negl. Trop. Dis. 6: e1892. doi:10.1371/journal.pntd.0001892.
- ↑ Hussain, M; Lu G; Torres S; Edmonds JH. (2013). "Effect of Wolbachia on replication of West Nile virus in a mosquito cell line and adult mosquitoes". J Virol. 87: 851–858. doi:10.1128/JVI.01837-12.
- ↑ Dodson, BL; Grant H; Oluwatobi P. (2014). "Wolbachia Enhances West Nile Virus (WNV) Infection in the Mosquito Culex tarsalis". PLOS. 8: e2965. doi:10.1371/journal.pntd.0002965.
- ↑ Pan, X; Zhou G; Wu J; et al. (2012). "Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti". Proc Natl Acad Sci U S A. 109 (1): E23–31. doi:10.1073/pnas.1116932108. PMC 3252928. PMID 22123956.
- ↑ Bian, G; Joshi D; Dong Y; et al. (2013). "Wolbachia invades Anopheles stephensi populations and induces refractoriness to Plasmodium infection". Science. 340 (6133): 748–51. doi:10.1126/science.1236192. PMID 23661760.
- ↑ Jason Gale (4 February 2016). "The Best Weapon for Fighting Zika? More Mosquitoes". Bloomberg.com.
- ↑ http://www.cell.com/cell-host-microbe/fulltext/S1931-3128%2816%2930157-3
- ↑ http://www.cidrap.umn.edu/news-perspective/2016/10/wolbachia-efforts-ramp-fight-zika-brazil-colombia
Further reading
- Werren, J.H. (1997). "Biology of Wolbachia" (PDF). Annual Review of Entomology. 42: 587–609. doi:10.1146/annurev.ento.42.1.587. PMID 15012323.
- Klasson L, Westberg J, Sapountzis P, Näslund K, Lutnaes Y, Darby AC, Veneti Z, Chen L, Braig HR, Garrett R, Bourtzis K, Andersson SG; Westberg; Sapountzis; Näslund; Lutnaes; Darby; Veneti; Chen; Braig; Garrett; Bourtzis; Andersson (23 March 2009). "The mosaic genome structure of the Wolbachia wRi strain infecting Drosophila simulans" (PDF). Proc. Natl. Acad. Sci. U.S.A. 106 (14): 5725–30. Bibcode:2009PNAS..106.5725K. doi:10.1073/pnas.0810753106. PMC 2659715. PMID 19307581.
External links
- Virtual Museum of Bacteria
- Wolbachia research portal National Science Foundation
- "One Species' Genome Discovered Inside Another's—Bacterial to Animal Gene Transfers Now Shown to be Widespread, with Implications for Evolution and Control of Diseases and Pests". University of Rochester. 30 August 2007. Retrieved 27 November 2007.
- Wolbachia at the Encyclopedia of Life
- Howard Hughes Medical Institute High School Lab Series