Vaginal flora
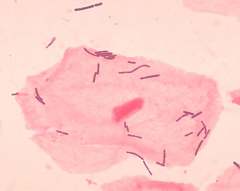
Vaginal flora or vaginal microbiota are the microorganisms that colonize the vagina. It was discovered by the German gynecologist Albert Döderlein in 1892.,[1] and are part of the overall human flora. The amount and type of bacteria present have significant implications for a woman's overall health. The primary colonizing bacteria of a healthy individual are of the genus Lactobacillus,[2] such as L. crispatus, and the lactic acid they produce is thought to protect against infection by pathogenic species.[3]
Lactobacilli
The primary colonizing bacteria of a healthy individual are of the genus Lactobacillus.[2] Since the first description of lactobacilli by Döderlein, lactobacilli have been generally considered as the gatekeepers of the vaginal ecosystem. Lactobacilli have been shown to inhibit in vitro growth of pathogenic microorganisms, e.g. Bacteroides fragilis, Escherichia coli, Gardnerella vaginalis, Mobiluncus spp., Neisseria gonorrhoeae, Peptostreptococcus anaerobius, P. bivia and Staphylococcus aureus. It is generally accepted that this is achieved mainly through the action of lactic acid.[4][5][6][7] Moreover, lactobacilli normally help to prevent long-term colonization of the vagina by adhering to vaginal epithelial cells. This usually reduces pathogens from infecting to the vaginal epithelium.[8]
Next to lactic acid production and competition for adherence, other antagonistic mechanisms include hydrogen peroxide (a broad-spectrum antimicrobial) and bacteriocins (target-specific antimicrobials) production.[9][10]
The production of lactic acid
Low pH is generally accepted to be the main mechanism controlling the composition of the vaginal microflora. Although the lactic acid produced by lactobacilli contributes to the vaginal acidity, it is still not proven to be the primary source of low vaginal pH, but the fact remains that most lactobacilli thrive best at a pH < 4.5 .[11][12][13]
Hydrogen peroxide
Production of hydrogen peroxide (H2O2) is a well-known mechanism for bacterial antagonism,[14][15][16] inhibiting growth of microorganisms via direct interaction or via human myeloperoxidase.[17][18][19] Hydrogen peroxide-producing lactobacilli have been shown to inactivate HIV-1, herpes simplex virus type 2 (HSV-2), Trichomonas vaginalis, G. vaginalis, P. bivia and E. coli.[10][20] found that 96% of Lactobacillus species from a healthy vaginal ecosystem produced H2O2 (L. jensenii and L. vaginalis produce the highest levels of H2O2,[9][21] whereas only 6% of the lactobacilli recovered from women with BV produced H2O2.[17] In agreement with this, L. iners, most frequently associated with disturbed vaginal microflora,[22][23] is a poor producer of H2O2.[24][25] Vaginal colonization by H2O2-producing lactobacilli has been associated with a decrease in the occurrence of bacterial vaginosis (BV).[26] However, more recently O‘Hanlon et al.[27] demonstrated that cervicovaginal fluid and semen have a significant H2O2-blocking activity and they later [10] demonstrated that physiological concentrations of H2O2 below 100 μM fail to inactivate any of the 17 tested BV-associated bacteria, e.g. A. vaginae, G. vaginalis, Mobiluncus spp., P. bivia, Prevotella corporis, Mycoplasma hominis, even in the presence of human myeloperoxidase, known to increase the microbicidal activity of H2O2.[10] Only supraphysiologic concentrations of exogenous H2O2 (0.34% w/v, 100 mM) were sufficient to inactivate BV-associated bacteria at which concentration it more potently inactivated vaginal lactobacilli (L. crispatus, L. gasseri, L. iners and L. jensenii). A concentration of 100 mM H2O2 is approximately 50-fold higher than lactobacilli are capable of producing even under optimal aerobic, low-antioxidant conditions, and approximately 5,000-fold higher than the estimated H2O2 concentration in vivo. Even more remarkable, the addition of only 1% vaginal fluid blocked the microbicidal activity of 1 M H2O2. Possible explanations may be that cervicovaginal fluid and semen contain proteins, glycoproteins, polysaccharides, lipids, and other molecules with the potential to react with and inactivate H2O2. In addition, the vagina is hypoxic most of the time, whereas lactobacilli require oxygen to produce hydrogen peroxide. It is also remarkable that catalase, which provides bacteria protection against toxic H2O2, is absent in lactobacilli,[17][28] and as such they would be unprotected against their own H2O2 production. In contrast, under optimal anaerobic growth conditions, physiological concentrations of lactic acid inactivated the BV-associated pathogens without affecting the vaginal lactobacilli.[10][27] In summary, although the hydrogen peroxide production of lactobacilli has been considered as an important antimicrobial component, contributing to the colonization resistance provided by lactobacilli,[9][29] and although there seems to be a link between H2O2-producing lactobacilli and normal vaginal microflora, recent data do not support this role for H2O2.[10][27]
Bacteriocins
Vaginal lactobacilli produce antimicrobial peptides, i.e. bacteriocins such as lactocin 160 and crispasin.[11] with inhibitory activity ranging from narrow (closely related Lactobacillus species) to broad (diverse groups of bacteria, including G. vaginalis and P. bivia),[6] and bacteriocin-like substances, with a broader spectrum of activity than bacteriocins (e.g. a heat-resistant peptide produced by Lactobacillus salivarius subsp. salivarius CRL 1328). Several studies have indicated that the activity of bacteriocins is favored by low pH.
The inhibitory substances produced by vaginal Lactobacillus is a primary factor in protecting the vaginal microbiota, with organic acids, bacteriocins, and hydrogen peroxide. These act synergistically against infection by pathogens. Not all Lactobacillus spp. and not all strains within one Lactobacillus species exhibit all 3 mechanisms.[11] Lactobacillus species differ in premenopausal women, i.e. L. crispatus, L. jensenii, L. iners, L. gasseri (and possibly L. vaginalis), as assessed through cultivation-dependent and cultivation-independent techniques.[22][23][24][30] Vaginal lactobacilli have been shown to display a pronounced vaginotropism, and their pili act as ligands for attachment to receptors of vaginal epithelial cells. The limited number of Lactobacillus spp. found in the human vagina is remarkable, which leads to the possibility that there are host factors that select for specific organisms, that these species have unusual characteristics that allow them to successfully colonize the vagina, or both .[31] However, the vaginotropism, does not only apply to this selected group of lactobacilli that stand for a healthy vagina, but also for the bacterial species associated with BV.[32] The microbiota detected in the human genital and gut econiche do not appear to grow outside their host and probably are likely to rely on the close contact between parents and their children for transmission,[32] e.g. mother to neonate transmission of genital microflora, most probably also with gut microflora homogenously distributed over the baby‘s body including skin, the oral cavity, nasopharynx, and feces.[33]
Other microbiota
The normal vaginal microbiota that is dominated by lactobacilli can differ among some ethnic groups. Non-pathogenic vaginal species are part of the normal microbiota of some women.[22][34] Several studies have demonstrated that a significant proportion (7–33%) of healthy asymptomatic women (especially black and Hispanic women)[35] lack appreciable numbers of Lactobacillus species in the vagina,[31][36] and instead have a vaginal microbiota that consist of other lactic acid-producing bacteria, i.e. species from the genera Atopobium, Leptotrichia, Leuconostoc, Megasphaera, Pediococcus, Streptococcus and Weissella,[30][31][35] All ethnic populations have vaginal microflora communities containing lactic acid producing bacteria.[31][31][35] This implies that not all communities may be equally resilient, so that if the resilience of a vaginal community is low then transitory changes in the structure of these communities may occur more readily in response to disturbances of various kinds, including menses, sexual intercourse, douching and contraceptive practices. These differences in the structure and composition of microbial communities may underlie well-known differences in the susceptibility of women in these racial groups to BV and various vaginal infections‖.[35][37][38] Although it can be agreed that we should be cautious about which vaginal microflora should be considered as disturbed, and although (lactic) acid is indeed produced by many species in each of these communities, the remark by Mirmonsef et al.[39] that emphasis should be on vaginal pH and not on mere acid production in establishing normal vaginal microbiota may be justified. The pH further decreases during pregnancy.[40]
Other vaginal bacterial species
Other bacterial species are frequently found in the vagina, such as the Gram positive cocci: Atopobium vaginae, Peptostreptococcus spp., Staphylococcus spp., Streptococcus spp., and Bacteroides spp., Fusobacterium spp., Gardnerella vaginalis, Mobiluncus, Prevotella spp., and Gram-negative enteric organisms, such as Escherichia coli.[22][23] Mycoplasma and Ureaplasma are frequently found in the vagina. Some of the obligate and facultative anaerobic bacteria are associated with BV.[36]
Pregnancy
The effect of tampon use on vaginal flora is debated, but application of tampons appears not to significantly modify the balance of bacterial presence. Pregnancy alters the microbiota with a reduction in species/genus diversity.[41]
Disease prevention
A healthy vaginal microbiome aids in the prevention of bacterial vaginosis, yeast infections and other possible problems by maintaining an acidic pH (< 4.5) that is unfavourable for the growth of common pathogens, such as Gardnerella vaginalis. The lactobacilli present in a healthy vaginal microbiome also occupy the ecological niche that would otherwise be available for exploitation by pathogenic organisms. However, harmful bacteria or an imbalance in bacteria can lead to infection.
Bacterial vaginosis is associated with the presence of Gardnerella vaginalis and Peptostreptococcus anaerobius[42] and a decrease in the number of Lactobacillus species that comprise the healthy vaginal microbiota.[41][43][44][45]
See also
References
- ↑ David, M. (2006). "Albert und Gustav Döderlein - ein kritischer Blick auf zwei besondere Lebensläufe deutscher Ordinarien". Zentralblatt für Gynäkologie. 128 (2): 56–59. doi:10.1055/s-2006-921412. ISSN 0044-4197.
- 1 2 Vasquez, A.; Jakobsson, T.; Ahrne, S.; Forsum, U.; Molin, G. (2002). "Vaginal Lactobacillus Flora of Healthy Swedish Women". Journal of Clinical Microbiology. 40 (8): 2746–2749. doi:10.1128/JCM.40.8.2746-2749.2002. PMC 120688
 . PMID 12149323.
. PMID 12149323. - ↑ Witkin, S. S.; Linhares, I. M.; Giraldo, P. (2007). "Bacterial flora of the female genital tract: Function and immune regulation". Best Practice & Research Clinical Obstetrics & Gynaecology. 21 (3): 347–354. doi:10.1016/j.bpobgyn.2006.12.004. PMID 17215167.
- ↑ Graver, M., and J. Wade. 2011. The role of acidification in the inhibition of Neisseria gonorrhoeae by vaginal lactobacilli during anaerobic growth. Ann. Clin. Microbiol. Antimicrob. 10:8.
- ↑ Matu, M. N., G. O. Orinda, E. N. M. Njagi, C. R. Cohen, and E. A. Bukusi. 2010. In vitro inhibitory activity of human vaginal lactobacilli against pathogenic bacteria associated with bacterial vaginosis in Kenyan women. Anaerobe 16:210-215.
- 1 2 Skarin, A., and J. Sylwan. 1986. Vaginal Lactobacilli inhibiting growth of Gardnerella vaginalis, Mobiluncus and other bacterial species cultured from vaginal content of women with bacterial vaginosis. APMIS. 94:399-403.
- ↑ Strus, M., M. Malinowska, and P. B. Heczko. 2002. In vitro antagonistic effect of Lactobacillus on organisms associated with bacterial vaginosis. J. Reprod. Med. 47:41-46.
- ↑ Boris, S., and C. Barbes. 2000. Role played by lactobacilli in controlling the population of vaginal pathogens. Microb. Infect. 2:543-546.
- 1 2 3 Martin, R., and J. E. Suarez. 2010 Biosynthesis and degradation of H2O2 by vaginal lactobacilli" Appl. Environ. Microbiol. 76:400-405.
- 1 2 3 4 5 6 O'Hanlon, D., T. Moench, and R. Cone. 2011. In vaginal fluid, bacteria associated with bacterial vaginosis can be suppressed with lactic acid but not hydrogen peroxide. BMC Infect. Dis.11:200.
- 1 2 3 Aroutcheva A.; Gariti D.; Simon M.; Shott S.; Faro J.; Simoes J. A.; Gurguis A.; Faro S. (2001). "Defense factors of vaginal lactobacilli". Am. J. Obstet. Gynecol. 185: 375–379.
- ↑ Linhares, I. M., P. R. Summers, B. Larsen, P. C. Giraldo, and S. S. Witkin. 2011. Contemporary perspectives on vaginal pH and lactobacilli" Am. J. Obstet. Gynecol. 204:120.e1-120.e5.
- ↑ Redondo-Lopez V.; Cook R. L.; Sobel J. D. (1990). "Emerging role of lactobacilli in the control and maintenance of the vaginal bacterial microflora". Rev. Infect. Dis. 12: 856–872. doi:10.1093/clinids/12.5.856.
- ↑ Dahiya R. S.; Speck M. L. (1968). "Hydrogen peroxide formation by lactobacilli and its effect on Staphylococcus aureus". J. Dairy Sci. 51: 1568–1572. doi:10.3168/jds.s0022-0302(68)87232-7.
- ↑ Thompson R.; Johnson A. (1951). "The inhibitory action of saliva on the diphtheria Bacillus: Hydrogen peroxide, the inhibitory agent produced by salivary streptococci". J. Infect. Dis. 88: 81–85. doi:10.1093/infdis/88.1.81.
- ↑ Wheater D. M.; Hirsch A.; Mattick A. T. R. (1952). "Possible identity of lactobacillin with hydrogen peroxide produced by lactobacilli". Nature. 170: 623–624. doi:10.1038/170623a0.
- 1 2 3 Eschenbach D. A.; Davick P. R.; Williams B. L.; Klebanoff S. J.; Young-Smith K.; Critchlow C. M.; Holmes K. K. (1989). "Prevalence of hydrogen peroxide-producing Lactobacillus species in normal women and women with bacterial vaginosis". J. Clin. Microbiol. 27: 251–256.
- ↑ Hillier S. L.; Krohn M. A.; Klebanoff S. J.; Eschenbach D. A. (1992). "The relationship of hydrogen peroxide-producing lactobacilli to bacterial vaginosis and genital microflora in pregnant women". Obstet. Gynecol. 79: 369–373. doi:10.1097/00006250-199203000-00008.
- ↑ Klebanoff SJ (1970). "Peroxidase-mediated antimicrobial activity of rat uterine fluid". Gynecol Invest. 1: 21–30. doi:10.1159/000301903.
- ↑ Baeten J. M.; Hassan W. M.; Chohan V.; Richardson B. A.; Mandaliya K.; Ndinya-Achola J. O.; Jaoko W.; McClelland R. S. (2009). "Prospective study of correlates of vaginal Lactobacillus colonisation among high-risk HIV-1 seronegative women". Sex. Transm. Infect. 85: 348–353. doi:10.1136/sti.2008.035451.
- ↑ Wilks, M., R. Wiggins, A. Whiley, E. Hennessy, S. Warwick, H. Porter, A. Corfield, and M. Millar. 2004. Identification and H2O2 production of vaginal lactobacilli from pregnant women at high risk of preterm birth and relation with outcome. J. Clin. Microbiol. 42:713-717.
- 1 2 3 4 Verhelst, R., H. Verstraelen, G. Claeys, G. Verschraegen, L. Van Simaey, C. De Ganck, E. De Backer, M. Temmerman, and M. Vaneechoutte. 2005. Comparison between Gram stain and culture for the characterization of vaginal microflora: Definition of a distinct grade that resembles grade I microflora and revised categorization of grade I microflora. BMC Microbiol. 5:61.
- 1 2 3 De Backer, E., R. Verhelst, H. Verstraelen, M. A. Alqumber, J. P. Burton, J. R. Tagg, M. Temmerman, and M. Vaneechoutte. 2007. Quantitative determination by real-time PCR of four vaginal Lactobacillus species, Gardnerella vaginalis and Atopobium vaginae indicates an inverse relationship between L. gasseri and L. iners. BMC Microbiol. 7:115
- 1 2 Antonio M. A.; Hawes S. E.; Hillier S. L. (1999). "The identification of vaginal Lactobacillus species and the demographic and microbiologic characteristics of women colonized by these species". J. Infect. Dis. 180: 1950–1956. doi:10.1086/315109. PMID 10558952.
- ↑ Antonio M. A. D.; Rabe L. K.; Hillier S. L. (2005). "Colonization of the rectum by Lactobacillus species and decreased risk of bacterial vaginosis". J. Infect. Dis. 192: 394–398. doi:10.1086/430926.
- ↑ Hawes S. E.; Hillier S. L.; Benedetti J.; Stevens C. E.; Koutsky L. A.; Wolner-Hanssen P. L.; Holmes K. K. (1996). "Hydrogen peroxide-producing lactobacilli and acquisition of vaginal infections". J. Infect. Dis. 174: 1058–1063. doi:10.1093/infdis/174.5.1058.
- 1 2 3 O'Hanlon, D. E., B. R. Lanier, T. R. Moench, and R. A. Cone. 2010. Cervicovaginal fluid and semen block the microbicidal activity of hydrogen peroxide produced by vaginal lactobacilli. BMC Infect. Dis.10:120.
- ↑ Klebanoff S. J.; Hillier S. L.; Eschenbach D. A.; Waltersdorph A. M. (1991). "Control of the microbial flora of the vagina by H202-generating lactobacilli". J. Infect. Dis. 164: 94–100. doi:10.1093/infdis/164.1.94.
- ↑ Vallor A. C.; Antonio M. A.; Hawes S. E.; Hillier S. L. (2001). "Factors associated with acquisition of, or persistent colonization by, vaginal lactobacilli: Role of hydrogen peroxide production". J. Infect. Dis. 184: 1431–1436. doi:10.1086/324445.
- 1 2 Ravel, Jacques; Gajer, Pawel; Abdo, Zaid; Schneider, G. Maria; Koenig, Sara S. K.; McCulle, Stacey L.; Karlebach, Shara; Gorle, Reshma; Russell, Jennifer; Tacket, Carol O.; Brotman, Rebecca M.; Davis, Catherine C.; Ault, Kevin; Peralta, Ligia; Forney, Larry J. (15 March 2011). "Vaginal microbiome of reproductive-age women". Proceedings of the National Academy of Sciences. 108 (Supplement 1): 4680–4687. doi:10.1073/pnas.1002611107. ISSN 0027-8424. Retrieved 27 May 2016.
- 1 2 3 4 5 Zhou X.; Bent S. J.; Schneider M. G.; Davis C. C.; Islam M. R.; Forney L. J. (2004). "Characterization of vaginal microbial communities in adult healthy women using cultivation-independent methods". Microbiol. 150: 2565–2573. doi:10.1099/mic.0.26905-0.
- 1 2 Danielsson D.; Teigen P. K.; Moi H. (2011). "The genital econiche: Focus on microbiota and bacterial vaginosis". Ann. N. Y. Acad. Sci. 1230: 48–58. doi:10.1111/j.1749-6632.2011.06041.x. PMID 21824165.
- ↑ Dominguez-Bello M. G.; Costello E. K.; Contreras M.; Magris M.; Hidalgo G.; Fierer N.; Knight R. (2010). "Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns". Proc. Natl. Acad. Sci. USA. 107: 11971–11975. doi:10.1073/pnas.1002601107.
- ↑ Lopes , Santos Santiago G., Cools P., Verstraelen H., Trog M., Missine G., Aila N. El, Verhelst R., Tency I., Claeys G., Temmerman M., Vaneechoutte M. (2011). "Longitudinal study of the dynamics of vaginal microflora during two consecutive menstrual cycles". PLOS ONE. 6: e28180. doi:10.1371/journal.pone.0028180.
- 1 2 3 4 Zhou X.; Brown C. J.; Abdo Z.; Davis C. C.; Hansmann M. A.; Joyce P.; Foster J. A.; Forney L. J. (2007). "Differences in the composition of vaginal microbial communities found in healthy Caucasian and black women". ISME J. 1: 121–133. doi:10.1038/ismej.2007.12.
- 1 2 Hummelen R.; Fernandes A. D.; Macklaim J. M.; Dickson R. J.; Changalucha J.; Gloor G. B.; Reid G. (2010). "Deep sequencing of the vaginal microbiota of women with HIV". PLOS ONE. 5: e12078. doi:10.1371/journal.pone.0012078.
- ↑ Martin J. A.; Hamilton B. E.; Sutton P. D.; Ventura S. J.; Mathews T. J.; Kirmeyer S.; Osterman M. J. (2010). "Births: Final data for 2007". National vital statistics reports. 58: 1–85.
- ↑ Ness R. B.; Hillier S.; Richter H. E.; Soper D. E.; Stamm C.; Bass D. C.; Sweet R. L.; Rice P. (2003). "Can known risk factors explain racial differences in the occurrence of bacterial vaginosis?". J. Natl. Med. Assoc. 95: 201–212.
- ↑ Mirmonsef P.; Gilbert D.; Veazey R. S.; Wang J.; Kendrick S. R.; Spear G. T. (2012). "A comparison of lower genital tract glycogen and lactic acid levels in women and macaques: Implications for HIV and SIV susceptibility". AIDS Res. Hum. Retroviruses. 28: 76–81. doi:10.1089/aid.2011.0071.
- ↑ Hillier S. L.; Nugent R. P.; Eschenbach D. A.; Krohn M. A.; Gibbs R. S.; Martin D. H.; Cotch M. F.; Edelman R.; Pastorek J. G.; Rao A. V.; McNellis D.; Regan J. A.; Carey J. C.; Klebanoff M. A. (1995). "Association between bacterial vaginosis and preterm delivery of a lowbirth-weight infant". N. Engl. J. Med. 333: 1737–1742. doi:10.1056/nejm199512283332604.
- 1 2 Clark, Natalie; Tal, Reshef; Sharma, Harsha; Segars, James (2014). "Microbiota and Pelvic Inflammatory Disease". Seminars in Reproductive Medicine. 32 (01): 043–049. doi:10.1055/s-0033-1361822. ISSN 1526-8004. PMC 4148456. PMID 24390920.
- ↑ "Bacterial Vaginosis (BV): Condition Information". National Institute of Child Health and Human Development. 2013-05-21. Retrieved 3 March 2015.
- ↑ Nardis, C.; Mastromarino, P.; Mosca, L. (September 2013). "Vaginal microbiota and viral sexually transmitted diseases". Annali di Igiene. 25 (5): 443–56. doi:10.7416/ai.2013.1946. PMID 24048183.
- ↑ "What are the symptoms of bacterial vaginosis?". National Institute of Child Health and Human Development. 2013-05-21. Retrieved 22 May 2016.
- ↑ Ravel, J.; et al. (2010). "Colloquium Paper: Vaginal microbiome of reproductive-age women". Proceedings of the National Academy of Sciences. 108 (Supplement_1): 4680–4687. doi:10.1073/pnas.1002611107. PMC 3063603. PMID 20534435.