Stichodactyla toxin
| ShK domain-like | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Rainbow colored cartoon diagram (N-terminus = blue, C-terminus = red) of an NMR solution structure of the ShK toxin.[1] Sidechains of cysteine residues involved in disulfide linkages are displayed as sticks and the sulfur atoms in these links are colored yellow. | |||||||||
| Identifiers | |||||||||
| Symbol | ShK | ||||||||
| Pfam | PF01549 | ||||||||
| InterPro | IPR003582 | ||||||||
| SMART | SM00254 | ||||||||
| SCOP | 1roo | ||||||||
| SUPERFAMILY | 1roo | ||||||||
| TCDB | 8.B.14 | ||||||||
| OPM superfamily | 475 | ||||||||
| OPM protein | 2lg4 | ||||||||
| |||||||||
Stichodactyla toxin (ShK) is a peptide toxin that blocks the voltage-gated potassium channels: Kv1.1, Kv1.3, Kv1.6, Kv3.2 and KCa3.1.
Structure
ShK is a 35-residue basic peptide first discovered in the sea anemone Stichodactyla helianthus by Professor Olga Castaneda from the University of Havana, Cuba, and her collaborators in Sweden. The formula is C169H274N54O48S7.[2] It is cross-linked by three disulfide bridges: Cys3-Cys35, Cys12-Cys28, and Cys17-Cys32 (see figure below).[3][4] The amino acid sequence of the ShK toxin is Arg-Ser-Cys-Ile-Asp-Thr-Ile-Pro-Lys-Ser-Arg-Cys-Thr-Ala-Phe-Gln-Cys-Lys-His-Ser-Met-Lys-Tyr-Arg-Leu-Ser-Phe-Cys-Arg-Lys-Thr-Cys-Gly-Thr-Cys.[2] ShK is stabilized by three disulfide bridges and consists of two short α-helices comprising residues 14-19 and 21-24.[1] The N-terminal eight residues of ShK adopt an extended conformation, followed by a pair of interlocking turns that resemble a 310 helix, while its C-terminal Cys35 residue forms a nearly head-to-tail cyclic structure through a disulfide bond with Cys3.[5] Protein domains with structural resemblance to ShK have been described in 688 proteins, most of them from C. elegans (InterPro: IPR003582). The SMART database at the EMBL has a list of 688 proteins containing 1315 ShK-like sequences (http://smart.embl-heidelberg.de). Other proteins containing domains with similar structures include the cysteine-rich secretory protein snake toxins natrin, triflin, and stecrisp, the Toxocara canis mucins, secreted peptides from the dog hookworm Ancylostoma caninum, and the human proteins Tpx-1 and matrix metalloprotease 23 (MMP23).[6][7][8][9][10][11][12]
Target
ShK toxin blocks the K+ channels Kv1.1, Kv1.3, Kv1.6, Kv3.2 and KCa3.1,[13][14][15][16][17] The peptide binds to all four subunits in the Kv1.3 tetramer through its interaction with the shallow vestibule at the outer entrance of the ion conduction pathway.[13][14][18] The peptide's Lysine22 residue occludes the channel pore like a "cork in a bottle". This blocks the entrance to the pore.[19][20]
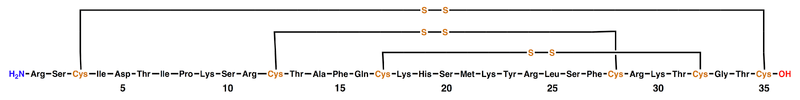
ShK blocks the Kv1.3 channel in T cells with a Kd of about 11 pM.[13][14][21] It blocks the neuronal Kv1.1 and Kv1.6 channels with Kds of 16 pM and 200 pM respectively.[16] The Kv3.2 and KCa3.1 channels are more than 1000 times less sensitive to the peptide.[13][14][16][17]
Several ShK analogs have been generated to enhance specificity for the Kv1.3 channel over the Kv1.1, Kv1.6 and Kv3.2 channels. The first analog that showed some degree of specificity was ShK-Dap22.[13] Attaching a fluorescein to the N-terminus of the peptide via a hydrophilic AEEA linker (2-aminoethoxy-2-ethoxy acetic acid; mini-PEG) resulted in a peptide, ShK-F6CA, with 100-fold specificity for Kv1.3 over Kv1.1 and related channels.[21] Based on this surprising finding additional analogs were made. ShK-170 [a.k.a. ShK(L5)],contains a L-phosphotyrosine in place of the fluorescein in ShK-F6CA. It blocks Kv1.3 with a Kd of 69 pM and shows exquisite specificity for Kv1.3.[16] However, it is chemically unstable. To improve stability a new analog, ShK-186 [a.k.a. SL5], was made with the C-terminal carboxyl of ShK-170 replaced by an amide; ShK-186 is otherwise identical to ShK-170.[22][23] In rats and squirrel monkeys, an indium-labeled ShK-186 analog called ShK-221, was slowly released from the injection site and maintained blood levels above the channel blocking dose for 3–5 days [24] ShK-192 is a new analog with increased stability.[23] It contains norleucine21 in place of methionine21 to avoid methionine oxidation, and the terminal phosphotyrosine is replaced by a non-hydrolyzable para-phosphonophenylalanine (Ppa) group.[23] ShK-192 is effective in ameliorating disease in rat models of multiple sclerosis. The D-diastereomer of ShK is also stable but blocks Kv1.3 with 2800-fold potency than the L-form (Kd = 36 nM) and it only exhibits 2-fold specificity for Kv1.3 over Kv1.1.[25] ShK-K-amide is a new analog with a C-terminal lysine. It blocks Kv1.3 with roughly 50-fold greater potency (IC50 of 26 ± 3 pM) than Kv1.1 ( IC50 of 942 ± 120 pM), and suppresses proliferation of human T cells (IC50 ≈ 3 nM).[26]
Kv1.3 and KCa3.1 regulate membrane potential and calcium signaling of T cells.[19] Calcium entry through the CRAC channel is promoted by potassium efflux through the Kv1.3 and KCa3.1 potassium channels.[22] Blockade of Kv1.3 channels in effector-memory T cells by ShK-186 suppresses calcium signaling, cytokine production (interferon-gamma, interleukin 2) and cell proliferation.[19][27][22] In vivo, ShK-186 paralyzes effector-memory T cells at the sites of inflammation and prevent their reactivation in inflamed tissues.[28] In contrast, ShK-186 does not affect the homing to and motility within lymph nodes of naive and central memory T cells, most likely because these cells express the KCa3.1 channel and are therefore protected from the effect of Kv1.3 blockade.[28] In proof-of-concept studies, ShK and its analogs have prevented and treated disease in rat models of multiple sclerosis, rheumatoid arthritis, and delayed type hypersensitivity.[21][29][16][29] ShK-186, due to its durable pharmacological action, is effective in ameliorating disease in rat models of delayed type hypersensitivity, multiple sclerosis (experimental autoimmune encephalomyelitis) and rheumatoid arthritis (pristane induced arthritis) when administered once every 2–5 days.[24] ShK-186 has completed non-clinical safety studies. ShK-186 is the subject of an open Investigational New Drug (IND) application in the USA, and has completed human phase 1A and 1B trials in healthy volunteers.
As ShK toxin binds to the synaptosomal membranes, it facilitates an acetylcholine release at avian neuromuscular junctions while the Kv3.2 channels are expressed in neurons that fire at a high frequency (such as cortical GABAergic interneurons), due to their fast activation and deactivation rates.[17] By blocking Kv3.2, ShK toxin depolarises the cortical GABAergic interneurons. Kv3.2 is also expressed in pancreatic beta cells. These cells are thought to play a role in their delayed-rectifier current, which regulates glucose-dependent firing. Therefore, ShK, as a Kv3.2 blocker, might be useful in the treatment of type-2 diabetes, although inhibition of the delayed-rectifier current has not yet been observed in human cells even when very high ShK concentrations were used.[17]
Toxicity
Toxicity of ShK toxin in mice is quite low. The median paralytic dose is about 25 mg/kg bodyweight (which translates to 0.5 mg per 20 g mouse). In rats the therapeutic safety index was greater than 75-fold.
ShK-Dap22 is less toxic, even a dose of 1.0 mg dose did not cause hyperactivity, seizures or mortality. The median paralytic dose was 200 mg/kg body weight.[13]
ShK-170 [a.k.a. ShK(L5)] does not cause significant toxicity in vitro. The peptide was not toxic to human and rat lymphoid cells incubated for 48 h with 100 nM of ShK-170 (>1200 times greater than the Kv1.3 half-blocking dose). The same high concentration of ShK-170 was negative in the Ames test on tester strain TA97A, suggesting that it is not a mutagen. ShK-170 had no effect on heart rate or heart rate variability parameters in either the time or the frequency domain in rats. It does not block the hERG (Kv11.1) channel that is associated with drug-associated cardiac arrhythmias. Repeated daily administration of the peptide by subcutaneous injection (10 µg/kg/day) for 2 weeks to rats does not cause any changes in blood counts, blood chemistry or in the proportion of thymocyte or lymphocyte subsets. Furthermore, the rats administered the peptide gain weight normally.
ShK-186 [a.k.a. SL5] is also safe. Repeated daily administration by subcutaneous injection of ShK-186 (100 µg/kg/day) for 4 weeks to rats does not cause any changes in blood counts, blood chemistry or histopathology.[22] Furthermore, ShK-186 did not compromise the protective immune response to acute influenza viral infection or acute bacterial (Chlamydia) infection in rats at concentrations that were effective in ameliorating autoimmune diseases in rat models.[28] Interestingly, rats repeatedly administered ShK-186 for a month by subcutaneous injection (500 µg/kg/day) developed low titer anti-ShK antibodies.[29] The reason for the low immunogenicity of the peptide is not well understood. ShK-186 has completed GLP (Good Laboratory Practice) non-clinical safety studies in rodents and non-human primates. ShK-186 (aka Dalazatide) which was licensed to Kineta Bio is the subject of an open Investigational New Drug (IND) application in the United States of America, and has recently completed human phase 1A and 1b trials in healthy volunteers. A second human phase 1b was recently completed in 2015 in psoriasis patients. Dalazatide was shown to significantly ameliorate symptoms in 90% patients with active plaque psoriasis with a 60 mcg weekly dose.
Many groups are developing Kv1.3 blockers for the treatment of autoimmune diseases.[30]
Use
Because ShK toxin is a specific inhibitor of Kv1.1, Kv1.3, Kv1.6, Kv3.2 and KCa3.1, it may serve as a useful pharmacological tool for studying these channels.[17][21] The Kv1.3 specific ShK analogs, ShK-170, ShK-186 and ShK-192, have been demonstrated to be effective in rat models of autoimmune diseases, and these or related analogs might have use as therapeutics for human autoimmune diseases.
Kv1.3 is also considered a therapeutic target for the treatment of obesity,[31][32] for enhancing peripheral insulin sensitivity in patients with type-2 diabetes mellitus,[33] and for preventing bone resorption in periodontal disease.[34] Furthermore, because pancreatic beta cells, which have Kv3.2 channels, are thought to play a role in glucose-dependent firing, ShK, as a Kv3.2 blocker, might be useful in the treatment of type-2 diabetes, although inhibition of the delayed-rectifier current has not yet been observed in human cells even when very high ShK concentrations were used.[16]
References
- 1 2 PDB: 1ROO; Tudor JE, Pallaghy PK, Pennington MW, Norton RS (April 1996). "Solution structure of ShK toxin, a novel potassium channel inhibitor from a sea anemone". Nat. Struct. Biol. 3 (4): 317–20. doi:10.1038/nsb0496-317. PMID 8599755.
- 1 2 Castañeda O, Sotolongo V, Amor AM, Stöcklin R, Anderson AJ, Harvey AL, Engström A, Wernstedt C, Karlsson E (May 1995). "Characterization of a potassium channel toxin from the Caribbean Sea anemone Stichodactyla helianthus". Toxicon. 33 (5): 603–13. doi:10.1016/0041-0101(95)00013-C. PMID 7660365.
- ↑ Pennington MW, Mahnir VM, Khaytin I, Zaydenberg I, Byrnes ME, Kem WR (December 1996). "An essential binding surface for ShK toxin interaction with rat brain potassium channels". Biochemistry. 35 (51): 16407–11. doi:10.1021/bi962463g. PMID 8987971.
- ↑ Pennington MW, Lanigan MD, Kalman K, Mahnir VM, Rauer H, McVaugh CT, Behm D, Donaldson D, Chandy KG, Kem WR, Norton RS (November 1999). "Role of disulfide bonds in the structure and potassium channel blocking activity of ShK toxin". Biochemistry. 38 (44): 14549–58. doi:10.1021/bi991282m. PMID 10545177.
- ↑ Pohl J, Hubalek F, Byrnes ME, Nielsen KR, Woods A, Pennington MW (1995). "Assignment of the three disulfide bonds in ShK toxin: A potent potassium channel inhibitor from the sea anemone Stichodactyla helianthus" (PDF). Letters in Peptide Science. 1 (6): 291–297. doi:10.1007/BF00119770.
- ↑ Wang F, Li H, Liu MN, Song H, Han HM, Wang QL, Yin CC, Zhou YC, Qi Z, Shu YY, Lin ZJ, Jiang T (December 2006). "Structural and functional analysis of natrin, a venom protein that targets various ion channels". Biochem. Biophys. Res. Commun. 351 (2): 443–8. doi:10.1016/j.bbrc.2006.10.067. PMID 17070778.
- ↑ Shikamoto Y, Suto K, Yamazaki Y, Morita T, Mizuno H (July 2005). "Crystal structure of a CRISP family Ca2+ -channel blocker derived from snake venom". J. Mol. Biol. 350 (4): 735–43. doi:10.1016/j.jmb.2005.05.020. PMID 15953617.
- ↑ Guo M, Teng M, Niu L, Liu Q, Huang Q, Hao Q (April 2005). "Crystal structure of the cysteine-rich secretory protein stecrisp reveals that the cysteine-rich domain has a K+ channel inhibitor-like fold". J. Biol. Chem. 280 (13): 12405–12. doi:10.1074/jbc.M413566200. PMID 15596436.
- ↑ Gibbs GM, Scanlon MJ, Swarbrick J, Curtis S, Gallant E, Dulhunty AF, O'Bryan MK (February 2006). "The cysteine-rich secretory protein domain of Tpx-1 is related to ion channel toxins and regulates ryanodine receptor Ca2+ signaling". J. Biol. Chem. 281 (7): 4156–63. doi:10.1074/jbc.M506849200. PMID 16339766.
- ↑ Loukas A, Hintz M, Linder D, Mullin NP, Parkinson J, Tetteh KK, Maizels RM (December 2000). "A family of secreted mucins from the parasitic nematode Toxocara canis bears diverse mucin domains but shares similar flanking six-cysteine repeat motifs". J. Biol. Chem. 275 (50): 39600–7. doi:10.1074/jbc.M005632200. PMID 10950959.
- ↑ Rangaraju S, Khoo KK, Feng ZP, Crossley G, Nugent D, Khaytin I, Chi V, Pham C, Calabresi P, Pennington MW, Norton RS, Chandy KG (March 2010). "Potassium channel modulation by a toxin domain in matrix metalloprotease 23". J Biol Chem. 285 (12): 9124–9136. doi:10.1074/jbc.M109.071266. PMC 2838332
 . PMID 19965868.
. PMID 19965868. - ↑ Loukas A, Prociv P (October 2001). "Immune responses in hookworm infections.". Clin Microbiol Rev. 14 (4): 689–703. doi:10.1128/CMR.14.4.689-703.2001. PMC 89000. PMID 11585781.
- 1 2 3 4 5 6 Kalman K, Pennington MW, Lanigan MD, Nguyen A, Rauer H, Mahnir V, Paschetto K, Kem WR, Grissmer S, Gutman GA, Christian EP, Cahalan MD, Norton RS, Chandy KG (December 1998). "ShK-Dap22, a potent Kv1.3-specific immunosuppressive polypeptide". J. Biol. Chem. 273 (49): 32697–707. doi:10.1074/jbc.273.49.32697. PMID 9830012.
- 1 2 3 4 Rauer H, Pennington M, Cahalan M, Chandy KG (July 1999). "Structural conservation of the pores of calcium-activated and voltage-gated potassium channels determined by a sea anemone toxin". J. Biol. Chem. 274 (31): 21885–92. doi:10.1074/jbc.274.31.21885. PMID 10419508.
- ↑ Middleton RE, Sanchez M, Linde AR, Bugianesi RM, Dai G, Felix JP, Koprak SL, Staruch MJ, Bruguera M, Cox R, Ghosh A, Hwang J, Jones S, Kohler M, Slaughter RS, McManus OB, Kaczorowski GJ, Garcia ML (November 2003). "Substitution of a single residue in Stichodactyla helianthus peptide, ShK-Dap22, reveals a novel pharmacological profile". Biochemistry. 42 (46): 13698–707. doi:10.1021/bi035209e. PMID 14622016.
- 1 2 3 4 5 6 Beeton C, Pennington MW, Wulff H, Singh S, Nugent D, Crossley G, Khaytin I, Calabresi PA, Chen CY, Gutman GA, Chandy KG (April 2005). "Targeting effector memory T cells with a selective peptide inhibitor of Kv1.3 channels for therapy of autoimmune diseases". Mol. Pharmacol. 67 (4): 1369–81. doi:10.1124/mol.104.008193. PMID 15665253.
- 1 2 3 4 5 Yan L, Herrington J, Goldberg E, Dulski PM, Bugianesi RM, Slaughter RS, Banerjee P, Brochu RM, Priest BT, Kaczorowski GJ, Rudy B, Garcia ML (May 2005). "Stichodactyla helianthus peptide, a pharmacological tool for studying Kv3.2 channels". Mol. Pharmacol. 67 (5): 1513–21. doi:10.1124/mol.105.011064. PMID 15709110.
- ↑ Lanigan MD, Kalman K, Lefievre Y, Pennington MW, Chandy KG, Norton RS (October 2002). "Mutating a critical lysine in ShK toxin alters its binding configuration in the pore-vestibule region of the voltage-gated potassium channel, Kv1.3". Biochemistry. 41 (40): 11963–71. doi:10.1021/bi026400b. PMID 12356296.
- 1 2 3 Chandy KG, Wulff H, Beeton C, Pennington M, Gutman GA, Cahalan MD (May 2004). "K+ channels as targets for specific immunomodulation". Trends Pharmacol. Sci. 25 (5): 280–9. doi:10.1016/j.tips.2004.03.010. PMC 2749963. PMID 15120495.
- ↑ Norton RS, Pennington MW, Wulff H (December 2004). "Potassium channel blockade by the sea anemone toxin ShK for the treatment of multiple sclerosis and othfer autoimmune diseases". Curr. Med. Chem. 11 (23): 3041–52. doi:10.2174/0929867043363947. PMID 15578998.
- 1 2 3 4 Beeton C, Wulff H, Singh S, Botsko S, Crossley G, Gutman GA, Cahalan MD, Pennington M, Chandy KG (March 2003). "A novel fluorescent toxin to detect and investigate Kv1.3 channel up-regulation in chronically activated T lymphocytes". J. Biol. Chem. 278 (11): 9928–37. doi:10.1074/jbc.M212868200. PMID 12511563.
- 1 2 3 4 Beeton C, Wulff H, Standifer NE, Azam P, Mullen KM, Pennington MW, Kolski-Andreaco A, Wei E, Grino A, Counts DR, Wang PH, LeeHealey CJ, S Andrews B, Sankaranarayanan A, Homerick D, Roeck WW, Tehranzadeh J, Stanhope KL, Zimin P, Havel PJ, Griffey S, Knaus HG, Nepom GT, Gutman GA, Calabresi PA, Chandy KG (November 2006). "Kv1.3 channels are a therapeutic target for T cell-mediated autoimmune diseases". Proc. Natl. Acad. Sci. U.S.A. 103 (46): 17414–9. doi:10.1073/pnas.0605136103. PMC 1859943. PMID 17088564.
- 1 2 3 Pennington MW, Beeton C, Galea CA, Smith BJ, Chi V, Monaghan KP, Garcia A, Rangaraju S, Giuffrida A, Plank D, Crossley G, Nugent D, Khaytin I, Lefievre Y, Peshenko I, Dixon C, Chauhan S, Orzel A, Inoue T, Hu X, Moore RV, Norton RS, Chandy KG (January 2009). "Engineering a stable and selective peptide blocker of the Kv1.3 channel in T lymphocytes". Mol. Pharmacol. 75 (4): 762–73. doi:10.1124/mol.108.052704. PMC 2684922. PMID 19122005.
- 1 2 Tarcha EJ, Chi V, Muñoz-Elias EJ, Bailey D, Londono LM, Upadhyay SK, Norton KN, Olson A, Tjong I, Nguyen HM, Hu X, Rupert GW, Boley SE, Slauter R, Sams J, Knapp B, Kentala D, Hansen Z, Pennington MW, Beeton C, Chandy KG, Iadonato SP (2012). "Durable pharmacological responses from a single dose of the peptide drug ShK-186, a specific Kv1.3 channel inhibitor". J. Pharm. Exp. Therap. 342 (3): 642–653. doi:10.1124/jpet.112.191890. PMID 22637724.
- ↑ Beeton C, Smith BJ, Sabo JK, Crossley G, Nugent D, Khaytin I, Chi V, Chandy KG, Pennington MW, Norton RS (January 2008). "The D-diastereomer of ShK toxin selectively blocks voltage-gated K+ channels and inhibits T lymphocyte proliferation". J. Biol. Chem. 283 (2): 988–97. doi:10.1074/jbc.M706008200. PMID 17984097.
- ↑ Pennington MW, Rashid MH, Tajhya RB, Beeton C, Kuyucak S, Norton RS (November 2012). "A C-terminally amidated analogue of ShK is a potent and selective blocker of the voltage-gated potassium channel Kv1.3". FEBS Lett. 586 (22): 3996–4001. doi:10.1016/j.febslet.2012.09.038. PMC 3496055. PMID 23063513.
- ↑ Wulff H, Calabresi PA, Allie R, Yun S, Pennington M, Beeton C, Chandy KG (June 2003). "The voltage-gated Kv1.3 K+ channel in effector memory T cells as new target for MS". J. Clin. Invest. 111 (11): 1703–13. doi:10.1172/JCI16921. PMC 156104. PMID 12782673.
- 1 2 3 Matheu MP, Beeton C, Garcia A, Chi V, Rangaraju S, Safrina O, Monaghan K, Uemura MI, Li D, Pal S, de la Maza LM, Monuki E, Flügel A, Pennington MW, Parker I, Chandy KG, Cahalan MD (October 2008). "Imaging of effector memory T cells during a delayed-type hypersensitivity reaction and suppression by Kv1.3 channel block". Immunity. 29 (4): 602–14. doi:10.1016/j.immuni.2008.07.015. PMC 2732399. PMID 18835197.
- 1 2 3 Beeton C, Wulff H, Barbaria J, Clot-Faybesse O, Pennington M, Bernard D, Cahalan MD, Chandy KG, Béraud E (November 2001). "Selective blockade of T lymphocyte K+ channels ameliorates experimental autoimmune encephalomyelitis, a model for multiple sclerosis". Proc. Natl. Acad. Sci. U.S.A. 98 (24): 13942–7. doi:10.1073/pnas.241497298. PMC 61146. PMID 11717451.
- ↑ Wulff H, Beeton C, Chandy KG (September 2003). "Potassium channels as therapeutic targets for autoimmune disorders". Curr Opin Drug Discov Devel. 6 (5): 640–7. PMID 14579513.
- ↑ Tucker K, Overton JM, Fadool DA (August 2008). "Kv1.3 gene-targeted deletion alters longevity and reduces adiposity by increasing locomotion and metabolism in melanocortin-4 receptor-null mice". Int J Obes (Lond). 32 (8): 1222–32. doi:10.1038/ijo.2008.77. PMC 2737548. PMID 18542083.
- ↑ Xu J, Koni PA, Wang P, Li G, Kaczmarek L, Wu Y, Li Y, Flavell RA, Desir GV (March 2003). "The voltage-gated potassium channel Kv1.3 regulates energy homeostasis and body weight". Hum. Mol. Genet. 12 (5): 551–9. doi:10.1093/hmg/ddg049. PMID 12588802.
- ↑ Xu J, Wang P, Li Y, Li G, Kaczmarek LK, Wu Y, Koni PA, Flavell RA, Desir GV (March 2004). "The voltage-gated potassium channel Kv1.3 regulates peripheral insulin sensitivity". Proc. Natl. Acad. Sci. U.S.A. 101 (9): 3112–7. doi:10.1073/pnas.0308450100. PMC 365752. PMID 14981264.
- ↑ Valverde P, Kawai T, Taubman MA (June 2005). "Potassium channel-blockers as therapeutic agents to interfere with bone resorption of periodontal disease". J. Dent. Res. 84 (6): 488–99. doi:10.1177/154405910508400603. PMID 15914584.