Single-domain antibody
.png)
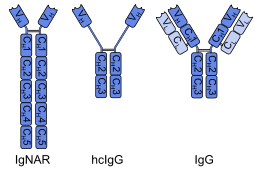
A single-domain antibody (sdAb, called Nanobody by Ablynx, the developer[1]) is an antibody fragment consisting of a single monomeric variable antibody domain. Like a whole antibody, it is able to bind selectively to a specific antigen. With a molecular weight of only 12–15 kDa, single-domain antibodies are much smaller than common antibodies (150–160 kDa) which are composed of two heavy protein chains and two light chains, and even smaller than Fab fragments (~50 kDa, one light chain and half a heavy chain) and single-chain variable fragments (~25 kDa, two variable domains, one from a light and one from a heavy chain).[2]
The first single-domain antibodies were engineered from heavy-chain antibodies found in camelids; these are called VHH fragments. Cartilaginous fishes also have heavy-chain antibodies (IgNAR, 'immunoglobulin new antigen receptor'), from which single-domain antibodies called VNAR fragments can be obtained. An alternative approach is to split the dimeric variable domains from common immunoglobulin G (IgG) from humans or mice into monomers. Although most research into single-domain antibodies is currently based on heavy chain variable domains, Nanobodies derived from light chains have also been shown to bind specifically to target epitopes.[3]
Single-domain camelids antibodies have been shown to be just as specific as a regular antibody and in some cases they are more robust. As well, they are easily isolated using the same phage panning procedure used for traditional antibodies, allowing them to be cultured in vitro in large concentrations. The smaller size and single domain make these antibodies easier to transform into bacterial cells for bulk production, making them ideal for research purposes. [4]
Single-domain antibodies are being researched for multiple pharmaceutical applications and have potential for use in the treatment of acute coronary syndrome, cancer and Alzheimer's disease.[5][6]
Properties
A single-domain antibody is a peptide chain of about 110 amino acids long, comprising one variable domain (VH) of a heavy-chain antibody, or of a common IgG. These peptides have similar affinity to antigens as whole antibodies, but are more heat-resistant and stable towards detergents and high concentrations of urea. Those derived from camelid and fish antibodies are less lipophilic and more soluble in water, owing to their complementarity determining region 3 (CDR3), which forms an extended loop (coloured orange in the ribbon diagram above) covering the lipophilic site that normally binds to a light chain.[7][8] In contrast to common antibodies, two out of six single-domain antibodies survived a temperature of 90 °C (194 °F) without losing their ability to bind antigens in a 1999 study.[9] Stability towards gastric acid and proteases depends on the amino acid sequence. Some species have been shown to be active in the intestine after oral application,[10][11] but their low absorption from the gut impedes the development of systemically active orally administered single-domain antibodies.
The comparatively low molecular mass leads to a better permeability in tissues, and to a short plasma half-life since they are eliminated renally.[2] Unlike whole antibodies, they do not show complement system triggered cytotoxicity because they lack an Fc region. Camelid and fish derived sdAbs are able to bind to hidden antigens that are not accessible to whole antibodies, for example to the active sites of enzymes. This property has been shown to result from their extended CDR3 loop, which is able to penetrate such sites.[8][12]
Production
From heavy-chain antibodies
A single-domain antibody can be obtained by immunization of dromedaries, camels, llamas, alpacas or sharks with the desired antigen and subsequent isolation of the mRNA coding for heavy-chain antibodies. By reverse transcription and polymerase chain reaction, a gene library of single-domain antibodies containing several million clones is produced. Screening techniques like phage display and ribosome display help to identify the clones binding the antigen.[13]
A different method uses gene libraries from animals that have not been immunized beforehand. Such naïve libraries usually contain only antibodies with low affinity to the desired antigen, making it necessary to apply affinity maturation by random mutagenesis as an additional step.[14]
When the most potent clones have been identified, their DNA sequence is optimized, for example to improve their stability towards enzymes. Another goal is humanization to prevent immunological reactions of the human organism against the antibody. Humanization is unproblematic because of the homology between camelid VHH and human VH fragments.[14] The final step is the translation of the optimised single-domain antibody in E. coli, Saccharomyces cerevisiae or other suitable organisms.
From conventional antibodies
Alternatively, single-domain antibodies can be made from common murine or human IgG with four chains.[15] The process is similar, comprising gene libraries from immunized or naïve donors and display techniques for identification of the most specific antigens. A problem with this approach is that the binding region of common IgG consists of two domains (VH and VL), which tend to dimerize or aggregate because of their lipophilicity. Monomerization is usually accomplished by replacing lipophilic by hydrophilic amino acids, but often results in a loss of affinity to the antigen.[16] If affinity can be retained, the single-domain antibodies can likewise be produced in E. coli, S. cerevisiae or other organisms.
Potential applications
Single-domain antibodies allow a broad range of applications in biotechnical as well as therapeutic use due to their small size, simple production and high affinity.[17]
Biotechnological and diagnostic
The fusion of a fluorescent protein to a nanobody generates a so-called chromobody. Chromobodies can be used to recognize and trace targets in different compartments of living cells. They can therefore increase the possibilities of live cell microscopy and will enable novel functional studies.[18] The coupling of an anti-GFP Nanobody to a monovalent matrix, called GFP-nanotrap, allows the isolation of GFP-fusion proteins and their interacting partners for further biochemical analyses.[19] Single molecule localization with super-resolution imaging techniques requires the specific delivery of fluorophores into close proximity with a target protein. Due to their large size the use of antibodies coupled to organic dyes can often lead to a misleading signal owing to the distance between the fluorophore and the target protein. The fusion of organic dyes to anti-GFP Nanobodies targeting GFP-tagged proteins allows nanometer spatial resolution and minimal linkage error because of the small size and high affinity.[20] To increase the crystallization probability of a target molecule, nanobodies can be used as crystallization chaperones. As auxiliary proteins, they can reduce the conformational heterogeneity by binding and stabilizing just a subset of conformational states. They also can mask surfaces interfering with the crystallization while extending regions that form crystal contacts.[21]
In diagnostic biosensor applications Nanobodies may be used prospectively as a tool. Due to their small size, they can be coupled more densely on biosensor surfaces. In addition to their advantage in targeting less accessible epitopes, their conformational stability also leads to higher resistance to surface regeneration conditions. After immobilizing single-domain antibodies on sensor surfaces sensing human prostate-specific antigen (hPSA) were tested. The Nanobodies outperformed the classical antibodies in detecting clinical significant concentrations of hPSA.[22]
Therapeutic
Single-domain antibodies have been tested as a new therapeutic tool against multiple targets. In mice infected with influenza A virus subtype H5N1, Nanobodies directed against hemaglutinin suppressed replication of the H5N1 virus in vivo and reduced morbidity and mortality.[23] Nanobodies targeting the cell receptor binding domain of the virulence factors toxin A and toxin B of Clostridium difficile were shown to neutralize cytopathic effects in fibroblasts in vitro.[24]
Orally available single-domain antibodies against E. coli-induced diarrhoea in piglets have been developed and successfully tested.[11] Other diseases of the gastrointestinal tract, such as inflammatory bowel disease and colon cancer, are also possible targets for orally available single-domain antibodies.[25]
Detergent-stable species targeting a surface protein of Malassezia furfur have been engineered for use in anti-dandruff shampoos.[7]

As an approach for photothermal therapy Nanobodies binding to the HER2 antigen, which is overexpressed in breast and ovarian cancer cells, were conjugated to branched gold nanoparticles (see figure). Tumor cells were destroyed photothermally using a laser in a test environment.[26]
ALX-0081, a single-domain antibody targeting von Willebrand factor is in clinical trials for the prevention of thrombosis in patients with acute coronary syndrome.[27] A Phase II study examining ALX-0081 in high risk percutaneous coronary intervention has started in September 2009.[28]
Ablynx expects that their Nanobodies might cross the blood–brain barrier and permeate into large solid tumours more easily than whole antibodies, which would allow for the development of drugs against brain cancers.[25]
References
- ↑ Gibbs, W. Wayt (August 2005). "Nanobodies". Scientific American Magazine.
- 1 2 Harmsen MM, De Haard HJ (November 2007). "Properties, production, and applications of camelid single-domain antibody fragments". Appl. Microbiol. Biotechnol. 77 (1): 13–22. doi:10.1007/s00253-007-1142-2. PMC 2039825
 . PMID 17704915.
. PMID 17704915. - ↑ Möller, A.; Pion, E; Narayan, V; Ball, KL (September 2010). "Intracellular activation of interferon regulatory factor-1 by nanobodies to the multi-functional (Mf1) domain". The Journal of Biological Chemistry. J Biol Chem. 285 (49): 38348–38361. doi:10.1074/jbc.M110.149476. PMC 2992268. PMID 20817723.
- ↑ Ghannam, A., Kumari, S., Muyldermans, S., & Abbady, A. Q. (2015). Camelid nanobodies with high affinity for broad bean mottle virus: a possible promising tool to immunomodulate plant resistance against viruses. Plant Molecular Biology, 1-15.
- ↑ "Nanobodies herald a new era in cancer therapy". The Medical News. 12 May 2004.
- ↑ "Pipeline". Ablynx. Retrieved 20 January 2010.
- 1 2 Dolk, E.; Van Der Vaart, M.; Lutje Hulsik, D.; Vriend, G.; De Haard, H.; Spinelli, S.; Cambillau, C.; Frenken, L.; Verrips, T. (2005). "Isolation of Llama Antibody Fragments for Prevention of Dandruff by Phage Display in Shampoo". Applied and Environmental Microbiology. 71 (1): 442–450. doi:10.1128/AEM.71.1.442-450.2005. PMC 544197. PMID 15640220.
- 1 2 Stanfield, R.; Dooley, H.; Flajnik, M.; Wilson, I. (2004). "Crystal structure of a shark single-domain antibody V region in complex with lysozyme". Science. 305 (5691): 1770–1773. Bibcode:2004Sci...305.1770S. doi:10.1126/science.1101148. PMID 15319492.
- ↑ Van Der Linden, R.; Frenken, L.; De Geus, B.; Harmsen, M.; Ruuls, R.; Stok, W.; De Ron, L.; Wilson, S.; Davis, P.; Verrips, C. T. (1999). "Comparison of physical chemical properties of llama VHH antibody fragments and mouse monoclonal antibodies". Biochimica et Biophysica Acta. 1431 (1): 37–46. doi:10.1016/S0167-4838(99)00030-8. PMID 10209277.
- ↑ Harmsen, M. .; Vansolt, C. .; Hoogendoorn, A. .; Vanzijderveld, F. .; Niewold, T. .; Vandermeulen, J. . (2005). "Escherichia coli F4 fimbriae specific llama single-domain antibody fragments effectively inhibit bacterial adhesion in vitro but poorly protect against diarrhoea". Veterinary Microbiology. 111 (1–2): 89–98. doi:10.1016/j.vetmic.2005.09.005. PMID 16221532.
- 1 2 Harmsen, M. M.; Van Solt, C. B.; Van Zijderveld-Van Bemmel, A. M.; Niewold, T. A.; Van Zijderveld, F. G. (2006). "Selection and optimization of proteolytically stable llama single-domain antibody fragments for oral immunotherapy". Applied Microbiology and Biotechnology. 72 (3): 544–551. doi:10.1007/s00253-005-0300-7. PMID 16450109.
- ↑ Desmyter, A.; Transue, T. R.; Ghahroudi, M. A.; Thi, M. H.; Poortmans, F.; Hamers, R.; Muyldermans, S.; Wyns, L. (1996). "Crystal structure of a camel single-domain VH antibody fragment in complex with lysozyme". Nature Structural Biology. 3 (9): 803–811. doi:10.1038/nsb0996-803. PMID 8784355.
- ↑ Arbabi Ghahroudi, M.; Desmyter, A.; Wyns, L.; Hamers, R.; Muyldermans, S. (1997). "Selection and identification of single domain antibody fragments from camel heavy-chain antibodies". FEBS Letters. 414 (3): 521–526. doi:10.1016/S0014-5793(97)01062-4. PMID 9323027.
- 1 2 Saerens, D.; Ghassabeh, G.; Muyldermans, S. (2008). "Single-domain antibodies as building blocks for novel therapeutics". Current Opinion in Pharmacology. 8 (5): 600–608. doi:10.1016/j.coph.2008.07.006. PMID 18691671.
- ↑ Holt, L. J.; Herring, C.; Jespers, L. S.; Woolven, B. P.; Tomlinson, I. M. (2003). "Domain antibodies: proteins for therapy". Trends in Biotechnology. 21 (11): 484–490. doi:10.1016/j.tibtech.2003.08.007. PMID 14573361.
- ↑ Borrebaeck, C. A. K.; Ohlin, M. (2002). "Antibody evolution beyond Nature". Nature Biotechnology. 20 (12): 1189–90. doi:10.1038/nbt1202-1189. PMID 12454662.
- ↑ Muyldermans, S. (2013). "Nanobodies: Natural Single-Domain Antibodies". Annual Review of Biochemistry. 82: 775–797. doi:10.1146/annurev-biochem-063011-092449. PMID 23495938.
- ↑ Rothbauer, U.; Zolghadr, K.; Tillib, S.; Nowak, D.; Schermelleh, L.; Gahl, A.; Backmann, N.; Conrath, K.; Muyldermans, S.; Cardoso, M. C.; Leonhardt, H. (2006). "Targeting and tracing antigens in live cells with fluorescent nanobodies". Nature Methods. 3 (11): 887–889. doi:10.1038/nmeth953. PMID 17060912.
- ↑ Rothbauer, U.; Zolghadr, K.; Muyldermans, S.; Schepers, A.; Cardoso, M. C.; Leonhardt, H. (2007). "A Versatile Nanotrap for Biochemical and Functional Studies with Fluorescent Fusion Proteins". Molecular & Cellular Proteomics. 7 (2): 282–289. doi:10.1074/mcp.M700342-MCP200. PMID 17951627.
- ↑ Ries, J.; Kaplan, C.; Platonova, E.; Eghlidi, H.; Ewers, H. (2012). "A simple, versatile method for GFP-based super-resolution microscopy via nanobodies". Nature Methods. 9 (6): 582–584. doi:10.1038/nmeth.1991. PMID 22543348.
- ↑ Koide, S. (2009). "Engineering of recombinant crystallization chaperones". Current Opinion in Structural Biology. 19 (4): 449–457. doi:10.1016/j.sbi.2009.04.008. PMC 2736338. PMID 19477632.
- ↑ Saerens, D.; Frederix, F.; Reekmans, G.; Conrath, K.; Jans, K.; Brys, L.; Huang, L.; Bosmans, E. N.; Maes, G.; Borghs, G.; Muyldermans, S. (2005). "Engineering Camel Single-Domain Antibodies and Immobilization Chemistry for Human Prostate-Specific Antigen Sensing". Analytical Chemistry. 77 (23): 7547–7555. doi:10.1021/ac051092j. PMID 16316161.
- ↑ Ibanez, L. I.; De Filette, M.; Hultberg, A.; Verrips, T.; Temperton, N.; Weiss, R. A.; Vandevelde, W.; Schepens, B.; Vanlandschoot, P.; Saelens, X. (2011). "Nanobodies with in Vitro Neutralizing Activity Protect Mice Against H5N1 Influenza Virus Infection". Journal of Infectious Diseases. 203 (8): 1063–1072. doi:10.1093/infdis/jiq168. PMID 21450996.
- ↑ Hussack, G.; Arbabi-Ghahroudi, M.; Van Faassen, H.; Songer, J. G.; Ng, K. K. - S.; MacKenzie, R.; Tanha, J. (2011). "Neutralization of Clostridium difficile Toxin a with Single-domain Antibodies Targeting the Cell Receptor Binding Domain". Journal of Biological Chemistry. 286 (11): 8961–8976. doi:10.1074/jbc.M110.198754. PMC 3058971. PMID 21216961.
- 1 2 "Nanobodies". Nanobody.org. 2006.
- ↑ Van De Broek, B.; Devoogdt, N.; d'Hollander, A.; Gijs, H. L.; Jans, K.; Lagae, L.; Muyldermans, S.; Maes, G.; Borghs, G. (2011). "Specific Cell Targeting with Nanobody Conjugated Branched Gold Nanoparticles for Photothermal Therapy". ACS Nano. 5 (6): 4319–4328. doi:10.1021/nn1023363. PMID 21609027.
- ↑ "Ablynx Announces Interim Results of First Nanobody Phase I Study of,ALX-0081 (ANTI-VWF)". Bio-Medicine.org. 2 July 2007.
- ↑ Clinical trial number NCT01020383 for "Comparative Study of ALX-0081 Versus GPIIb/IIIa Inhibitor in High Risk Percutaneous Coronary Intervention (PCI) Patients" at ClinicalTrials.gov
Engineered monoclonal antibodies and antibody mimetics | ||
|---|---|---|
| Whole antibody | 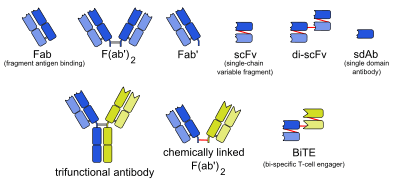 | |
| Fab fragment | ||
| Variable fragment | ||
| Smaller units | ||
| Intracellular | ||
| Antibody mimetics | ||