Short-term effects of alcohol consumption
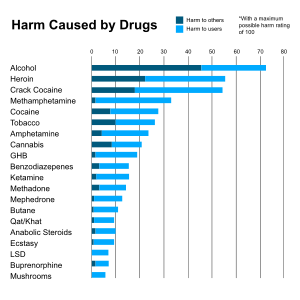
The short-term effects of alcohol (ethanol) consumption–due to drinking beer, wine, distilled spirits or other alcoholic beverages–range from a decrease in anxiety and motor skills and euphoria at lower doses to intoxication (drunkenness), stupor, unconsciousness, anterograde amnesia (memory "blackouts"), and central nervous system depression at higher doses. Cell membranes are highly permeable to alcohol, so once alcohol is in the bloodstream it can diffuse into nearly every cell in the body.
The concentration of alcohol in blood is measured via blood alcohol content (BAC). The amount and circumstances of consumption play a large part in determining the extent of intoxication; for example, eating a heavy meal before alcohol consumption causes alcohol to absorb more slowly.[1] The amount of alcohol consumed largely determines the extent of hangovers, although hydration also plays a role. After excessive drinking, stupor and unconsciousness can occur. Extreme levels of consumption can lead to alcohol poisoning and death (a concentration in the blood stream of 0.40% will kill half of those affected[2][3]). Alcohol may also cause death indirectly, by asphyxiation from vomit.
Alcohol can greatly exacerbate sleep problems. During abstinence, residual disruptions in sleep regularity and sleep patterns are the greatest predictors of relapse.[4]
Effects by dosage
Different concentrations of alcohol in the human body have different effects on the subject.
The following lists the common effects of alcohol on the body, depending on the blood alcohol concentration (BAC). However, tolerance varies considerably between individuals, as does individual response to a given dosage; the effects of alcohol differ widely between people. Hence, BAC percentages are just estimates used for illustrative purposes.
|
|
Moderate doses
Ethanol inhibits the ability of glutamate to open the cation channel associated with the N-methyl-D-aspartate (NMDA) subtype of glutamate receptors. Stimulated areas include the cortex, hippocampus and nucleus accumbens, which are responsible for thinking and pleasure seeking. Another one of alcohol's agreeable effects is body relaxation, possibly caused by neurons transmitting electrical signals in an alpha waves-pattern; such waves are observed (with the aid of EEGs) when the body is relaxed.
Short-term effects of alcohol include the risk of injuries, violence and fetal damage.[5] Alcohol has also been linked with lowered inhibitions, though it is unclear to what degree this is chemical versus psychological as studies with placebos can often duplicate the social effects of alcohol at low to moderate doses. Some studies have suggested that intoxicated people have much greater control over their behavior than is generally recognized, though they have a reduced ability to evaluate the consequences of their behavior.[6] Behavioral changes associated with drunkenness are, to some degree, contextual.[7][8]
Areas of the brain responsible for planning and motor learning are sharpened. A related effect, caused by even low levels of alcohol, is the tendency for people to become more animated in speech and movement. This is due to increased metabolism in areas of the brain associated with movement, such as the nigrostriatal pathway. This causes reward systems in the brain to become more active, which may induce certain individuals to behave in an uncharacteristically loud and cheerful manner.
Alcohol has been known to mitigate the production of antidiuretic hormone, which is a hormone that acts on the kidney to favour water reabsorption in the kidneys during filtration. This occurs because alcohol confuses osmoreceptors in the hypothalamus, which relay osmotic pressure information to the posterior pituitary, the site of antidiuretic hormone release. Alcohol causes the osmoreceptors to signal that there is low osmotic pressure in the blood, which triggers an inhibition of the antidiuretic hormone. As a consequence, one's kidneys are no longer able to reabsorb as much water as they should be absorbing, leading to creation of excessive volumes of urine and the subsequent overall dehydration.
Excessive doses
Acute alcohol intoxication through excessive doses in general causes short- or long-term health effects. NMDA receptors start to become unresponsive, slowing areas of the brain for which they are responsible. Contributing to this effect is the activity that alcohol induces in the gamma-aminobutyric acid (GABA) system. The GABA system is known to inhibit activity in the brain. GABA could also be responsible for the memory impairment that many people experience. It has been asserted that GABA signals interfere with the registration and consolidation stages of memory formation. As the GABA system is found in the hippocampus (among other areas in the CNS), which is thought to play a large role in memory formation, this is thought to be possible.
Anterograde amnesia, colloquially referred to as "blacking out", is another symptom of heavy drinking. This is the loss of memory during and after an episode of drinking. When alcohol is consumed at a rapid rate, the point at which most healthy people's long-term memory creation starts to fail usually occurs at approximately 0.20% BAC, but can be reached as low as 0.14% BAC for inexperienced drinkers.
Another classic finding of alcohol intoxication is ataxia, in its appendicular, gait, and truncal forms. Appendicular ataxia results in jerky, uncoordinated movements of the limbs, as though each muscle were working independently from the others. Truncal ataxia results in postural instability; gait instability is manifested as a disorderly, wide-based gait with inconsistent foot positioning. Ataxia is responsible for the observation that drunk people are clumsy, sway back and forth, and often fall down. It is presumed to be due to alcohol's effect on the cerebellum.
Allergic reaction-like symptoms
Humans metabolize ethanol primarily through NAD+-dependent alcohol dehydrogenase (ADH) class I enzymes (i.e. ADH1A, ADH1B, and ADH1C) to acetaldehyde and then metabolize acetaldehyde primarily by NAD2-dependent aldehyde dehydrogenase 2 (ALDH2) to acetic acid.[9][10] Eastern Asians reportedly have a deficiency in acetaldehyde metabolism in a surprisingly high percentage (approaching 50%) of their populations. The issue has been most thoroughly investigated in native Japanese where persons with a single-nucleotide polymorphism (SNP) variant allele of the ALDH2 gene were found; the variant allele, encodes lysine (lys) instead of glutamic acid (glu) at amino acid 487; this renders the enzyme essentially inactive in metabolizing acetaldehyde to acetic acid.[11][12] The variant allele is variously termed glu487lys, ALDH2*2, and ALDH2*504lys. In the overall Japanese population, about 57% of individuals are homozygous for the normal allele (sometimes termed ALDH2*1), 40% are heterozygous for glu487lys, and 3% are homozygous for glu487lys.[12] Since ALDH2 assembles and functions as a tetramer and since ALDH2 tetramers containing one or more glu487lys proteins are also essentially inactive (i.e. the variant allele behaves as a dominant negative), homozygote individuals for glu487lys have undetectable while heterozygote individuals for glu487lys have little ALDH2 activity.[13] In consequence, Japanese individuals homozygous or, to only a slightly lesser extent, homozygous for glu487lys metabolize ethanol to acetaldehyde normally but metabolize acetaldehyde poorly and are susceptible to a set of adverse responses to the ingestion of, and sometimes even the fumes from, ethanol and ethanol-containing beverages; these responses include the transient accumulation of acetaldehyde in blood and tissues; facial flushing (i.e. the "oriental flushing syndrome" or Alcohol flush reaction), urticaria, systemic dermatitis, and alcohol-induced respiratory reactions (i.e. rhinitis and, primarily in patients with a history of asthma, mild to moderately bronchoconstriction exacerbations of their asthmatic disease.[14] These allergic reaction-like symptoms, which typically occur within 30–60 minutes of ingesting alcoholic beverages, do not appear to reflect the operation of classical IgE- or T cell-related allergen-induced reactions but rather are due, at least in large part, to the action of acetaldehyde in stimulating tissues to release histamine, the probable evoker these symptoms.[14][15]
The percentages of glu487lys heterozygous plus homozygous genotypes are about 35% in native Caboclo of Brazil, 30% in Chinese, 28% in Koreans, 11% in Thai people, 7% in Malaysians, 3% in natives of India, 3% in Hungarians, and 1% in Filipinos; percentages are essentially 0 in individuals of Native African descent, Caucasians of Western European descent, Turks, Australian Aborigines, Australians of Western European descent, Swedish Lapps, and Alaskan Eskimos.[16][17] The prevalence of ethanol-induced allergic symptoms in 0 or low levels of glu487lys genotypes commonly ranges above 5%. These "ethanol reactors" may have other gene-based abnormalities that cause the accumulation of acetaldehyde following the ingestion of ethanol or ethanol-containing beverages. For example, the surveyed incidence of self-reported ethanol-induced flushing reactions in Scandinavians living in Copenhagen as well as Australians of European descent is about ~16% in individuals homozygous for the "normal" ADH1B gene but runs to ~23% in individuals with the ADH1-Arg48His SNP variant; in vitro, this variant metabolizes ethanol rapidly and in humans, it is proposed, may form acetaldehyde at levels that exceed the capacity of ALDH2 to metabolize the acetaldehyde.[17][18] Notwithstanding such considerations, experts suggest that the large proportion of alcoholic beverage -induced allergic-like symptoms in populations with a low incidence the glu487lys genotype reflect true allergic reactions to the natural and/or contaminating allergens particularly those in wines and to a lesser extent beers.[14]
Sleep
Moderate alcohol consumption and sleep disruptions
Moderate alcohol consumption 30–60 minutes before sleep, although decreasing, disrupts sleep architecture. Rebound effects occur once the alcohol has been largely metabolized, causing late night disruptions in sleep maintenance. Under conditions of moderate alcohol consumption where blood alcohol levels average 0.06–0.08 percent and decrease 0.01–0.02 percent per hour, an alcohol clearance rate of 4–5 hours would coincide with disruptions in sleep maintenance in the second half of an 8-hour sleep episode. In terms of sleep architecture, moderate doses of alcohol facilitate "rebounds" in rapid eye movement (REM) following suppression in REM and stage 1 sleep in the first half of an 8-hour sleep episode, REM and stage 1 sleep increase well beyond baseline in the second half. Moderate doses of alcohol also very quickly increase (SWS) in the first half of an 8-hour sleep episode. Enhancements in REM sleep and SWS following moderate alcohol consumption are mediated by reductions in glutamatergic activity by adenosine in the central nervous system. In addition, tolerance to changes in sleep maintenance and sleep architecture develops within 3 days of alcohol consumption before bedtime.
Alcohol consumption and sleep improvements
Low doses of alcohol (one 360 ml (13 imp fl oz; 12 US fl oz) beer) appear to increase total sleep time and reduce awakening during the night. The sleep-promoting benefits of alcohol dissipate at moderate and higher doses of alcohol.[19] Previous experience with alcohol also influences the extent to which alcohol positively or negatively affects sleep. Under free-choice conditions, in which subjects chose between drinking alcohol or water, inexperienced drinkers were sedated while experienced drinkers were stimulated following alcohol consumption.[20] In insomniacs, moderate doses of alcohol improve sleep maintenance.[21]
Alcohol consumption and fatigue
Conditions of fatigue correlate positively with increased alcohol consumption.
Alcohol abstinence and sleep disruptions
Hormonal imbalance and sleep disruptions following withdrawal from chronic alcohol consumption are strong predictors of relapse. During abstinence, recovering alcoholics have attenuated melatonin secretion at onset of a sleep episode, resulting in prolonged sleep onset latencies. Psychiatry and core body temperatures during the sleep period contribute to poor sleep maintenance. The effect of alcohol consumption on the circadian control of human core body temperature is time dependent.
Alcohol consumption and balance
Alcohol can affect balance by altering the viscosity of the endolymph within the otolithic membrane, the fluid inside the semicircular canals inside the ear. The endolymph surrounds the ampullary cupula which contains hair cells within the semicircular canals. When the head is tilted, the endolymph flows and moves the cupula. The hair cells then bend and send signals to the brain indicating the direction in which the head is tilted. By changing the viscosity of the endolymph to become less dense when alcohol enters the system, the hair cells can move more easily within the ear, which sends the signal to the brain and results in exaggerated and overcompensated movements of body. This can also result in vertigo, or "the spins."[22]
Pathophysiology
At low or moderate doses, alcohol acts primarily as a positive allosteric modulator of GABAA. Alcohol binds to several different subtypes of GABAA, but not to others. The main subtypes responsible for the subjective effects of alcohol are the α1β3γ2, α5β3γ2, α4β3δ and α6β3δ subtypes, although other subtypes such as α2β3γ2 and α3β3γ2 are also affected. Activation of these receptors causes most of the effects of alcohol such as relaxation and relief from anxiety, sedation, ataxia and increase in appetite and lowering of inhibitions that can cause a tendency toward violence in some people.[23][24][25][26][27][28][29]
Alcohol has a powerful effect on glutamate as well. Alcohol decreases glutamate's ability to bind with NMDA and acts as an antagonist of the NMDA receptor, which plays a critical role in LTP by allowing Ca2+ to enter the cell. These inhibitory effects are thought to be responsible for the "memory blanks" that can occur at levels as low as 0.03% blood level. In addition, reduced glutamate release in the dorsal hippocampus has been linked to spatial memory loss. Chronic alcohol users experience an upregulation of NMDA receptors because the brain is attempting to reestablish homeostasis. When a chronic alcohol user stops drinking for more than 10 hours, apoptosis can occur due to excitotoxicity. The seizures experienced during alcohol abstinence are thought to be a result of this NMDA upregulation. Alteration of NMDA receptor numbers in chronic alcoholics is likely to be responsible for some of the symptoms seen in delirium tremens during severe alcohol withdrawal, such as delirium and hallucinations. Other targets such as sodium channels can also be affected by high doses of alcohol, and alteration in the numbers of these channels in chronic alcoholics is likely to be responsible for as well as other effects such as cardiac arrhythmia. Other targets that are affected by alcohol include cannabinoid, opioid and dopamine receptors, although it is unclear whether alcohol affects these directly or if they are affected by downstream consequences of the GABA/NMDA effects. People with a family history of alcoholism may exhibit genetic differences in the response of their NMDA glutamate receptors as well as the ratios of GABAA subtypes in their brain.[30][31][32][33][34][35][36][37][38][39] Alcohol inhibits sodium-potassium pumps in the cerebellum and this is likely how it corrupts cerebellar computation and body co-ordination.[40][41]
Contrary to popular belief, research suggests that acute exposure to alcohol is not neurotoxic in adults and actually prevents NMDA antagonist-induced neurotoxicity.[42]
Research
Animal models using mammals and invertebrates have been informative in studying the effects of ethanol on not only pharmacokinetics of alcohol but also pharmacodynamics, in particular in the nervous system. Ethanol-induced intoxication is not uncommon in the animal kingdom, as noted here:
"Many of us have noticed that bees or yellow jackets cannot fly well after having drunk the juice of overripe fruits or berries; bears have been seen to stagger and fall down after eating fermented honey; and birds often crash or fly haphazardly while intoxicated on ethanol that occurs naturally as free-floating microorganisms convert vegetable carbohydrates to alcohol."[43]
More recently, studies using animal models have begun to elucidate the effects of ethanol on the nervous system. Traditionally, many studies have been performed in mammals, such as mice, rats, and non-human primates. However, non-mammalian animal models have also been employed; in particular, Ulrike Heberlein group at UC San Francisco has used Drosophila melanogaster, the fruit fly, taking advantage of its facile genetics to dissect the neural circuits and molecular pathways, upon which ethanol acts. The series of studies carried in the Heberlein lab has identified insulin and its related signaling pathways as well as biogenic amines in the invertebrate nervous system as being important in alcohol tolerance.[44][45][46] The value of antabuse (disulfiram) as a treatment for alcoholism has been tested using another invertebrate animal model, the honey bees.[47] It is important to note that some of the analogous biochemical pathways and neural systems have been known to be important in alcohol's effects on humans, while the possibility that others may also be important remains unknown. Research of alcohol's effects on the nervous system remains a hot topic of research, as scientists inch toward understanding the problem of alcohol addiction.
In addition to the studies carried out in invertebrates, researchers have also used vertebrate animal models to study various effects of ethanol on behaviors.
See also
References
- ↑ Horowitz M, Maddox A, Bochner M, et al. (August 1989). "Relationships between gastric emptying of solid and caloric liquid meals and alcohol absorption". Am. J. Physiol. 257 (2 Pt 1): G291–8. PMID 2764113.
- ↑ Alcohol Awareness Page
- ↑ Carleton College: Wellness Center: Blood Alcohol Concentration (BAC)
- ↑ Feige B, Scaal S, Hornyak M, Gann H, Riemann D (January 2007). "Sleep electroencephalographic spectral power after withdrawal from alcohol in alcohol-dependent patients". Alcohol. Clin. Exp. Res. 31 (1): 19–27. doi:10.1111/j.1530-0277.2006.00260.x. PMID 17207097.
- ↑ Andréasson, S.; Allebeck, P. (28 February – 6 March 2005). "[Alcohol as medication is no good. More risks than benefits according to a survey of current knowledge]". Lakartidningen. 102 (9): 632–7. PMID 15804034.
- ↑ Grattan KE, Vogel-Sprott M (February 2001). "Maintaining intentional control of behavior under alcohol". Alcohol. Clin. Exp. Res. 25 (2): 192–7. doi:10.1111/j.1530-0277.2001.tb02198.x. PMID 11236832.
- ↑ Grant NK, Macdonald TK (2005). "Can alcohol lead to inhibition or disinhibition? Applying alcohol myopia to animal experimentation". Alcohol Alcohol. 40 (5): 373–8. doi:10.1093/alcalc/agh177. PMID 15996970.
- ↑ http://www.sirc.org/publik/drinking4.html
- ↑ Ann Ist Super Sanita. 2006;42(1):8-16
- ↑ Proc Nutr Soc. 2004 Feb;63(1):49-63
- ↑ Science. 1972 Jan 28;175(4020):449–50
- 1 2 J Allergy Clin Immunol. 1998 May;101(5):576-80
- ↑ Pharmacol Rev. 2012 Jul;64(3):520-39. doi: 10.1124/pr.111.005538
- 1 2 3 Ann Allergy Asthma Immunol. 2013 Dec;111(6):439-45. doi: 10.1016/j.anai.2013.09.016
- ↑ Hum Mol Genet. 2009 Feb 1;18(3):580–93. doi: 10.1093/hmg/ddn372
- ↑ Hum Genet. 1992 Jan;88(3):344-6.
- 1 2 Hum Mol Genet. 2009 Feb 1;18(3):580–93. doi: 10.1093/hmg/ddn372>
- ↑ Int Arch Allergy Immunol. 2010;153(1):86-94. doi: 10.1159/000301583
- ↑ Stone BM (June 1980). "Sleep and low doses of alcohol". Electroencephalogr Clin Neurophysiol. 48 (6): 706–9. doi:10.1016/0013-4694(80)90427-7. PMID 6155259.
- ↑ Schuckit MA (February 1994). "Low level of response to alcohol as a predictor of future alcoholism". Am J Psychiatry. 151 (2): 184–9. PMID 8296886.
- ↑ Roehrs T, Papineau K, Rosenthal L, Roth T (March 1999). "Ethanol as a hypnotic in insomniacs: self administration and effects on sleep and mood". Neuropsychopharmacology. 20 (3): 279–86. doi:10.1016/S0893-133X(98)00068-2. PMID 10063488.
- ↑ , Why does my head spin when I'm drunk? Rachel Nuwer, 8 Feb 2011, Scienceline.
- ↑ Huang Q, He X, Ma C, et al. (January 2000). "Pharmacophore/receptor models for GABAA/BzR subtypes (α1β3γ2, α5β3γ2, and α6β3γ2) via a comprehensive ligand-mapping approach". J. Med. Chem. 43 (1): 71–95. doi:10.1021/jm990341r. PMID 10633039.
- ↑ Platt DM, Duggan A, Spealman RD, et al. (May 2005). "Contribution of α1GABAA and α5GABAA receptor subtypes to the discriminative stimulus effects of ethanol in squirrel monkeys". J. Pharmacol. Exp. Ther. 313 (2): 658–67. doi:10.1124/jpet.104.080275. PMID 15650112.
- ↑ Duke AN, Platt DM, Cook JM, et al. (August 2006). "Enhanced sucrose pellet consumption induced by benzodiazepine-type drugs in squirrel monkeys: role of GABAA receptor subtypes". Psychopharmacology (Berl.). 187 (3): 321–30. doi:10.1007/s00213-006-0431-2. PMID 16783540.
- ↑ Wallner M, Hanchar HJ, Olsen RW (May 2006). "Low-dose alcohol actions on α4β3δ GABAA receptors are reversed by the behavioral alcohol antagonist Ro15-4513". Proc. Natl. Acad. Sci. U.S.A. 103 (22): 8540–5. doi:10.1073/pnas.0600194103. PMC 1482527
 . PMID 16698930.
. PMID 16698930. - ↑ Mehta AK, Ticku MK (August 1988). "Ethanol potentiation of GABAergic transmission in cultured spinal cord neurons involves gamma-aminobutyric acidA-gated chloride channels". J. Pharmacol. Exp. Ther. 246 (2): 558–64. PMID 2457076.
- ↑ Becker HC, Anton RF (January 1989). "The benzodiazepine receptor inverse agonist RO15-4513 exacerbates, but does not precipitate, ethanol withdrawal in mice". Pharmacol. Biochem. Behav. 32 (1): 163–7. doi:10.1016/0091-3057(89)90227-X. PMID 2543989.
- ↑ Hanchar HJ, Chutsrinopkun P, Meera P, et al. (May 2006). "Ethanol potently and competitively inhibits binding of the alcohol antagonist Ro15-4513 to α4/6β3δ GABAA". Proc. Natl. Acad. Sci. U.S.A. 103 (22): 8546–51. doi:10.1073/pnas.0509903103. PMC 1482528. PMID 16581914.
- ↑ Shimizu, A. Matsubara, K. Uezono, T. Kimura, K. Shiono, H. "Reduced dorsal hippocampal glutamate release significantly correlates with the spatial memory deficits produced by benzodiazepines and ethanol". Neuroscience. P 701-706
- ↑ Meyer, Jerrold. Quenzer, Linda. "Psychopharmacology: drugs brain and behavior"
- ↑ Petrakis IL, Limoncelli D, Gueorguieva R, et al. (October 2004). "Altered NMDA glutamate receptor antagonist response in individuals with a family vulnerability to alcoholism". Am J Psychiatry. 161 (10): 1776–82. doi:10.1176/appi.ajp.161.10.1776. PMID 15465973.
- ↑ Nutt DJ (May 2006). "Alcohol alternatives—a goal for psychopharmacology?". J. Psychopharmacol. (Oxford). 20 (3): 318–20. doi:10.1177/0269881106063042. PMID 16574703.
- ↑ Qiang M, Denny AD, Ticku MK (July 2007). "Chronic intermittent ethanol treatment selectively alters N-methyl-D-aspartate receptor subunit surface expression in cultured cortical neurons". Mol. Pharmacol. 72 (1): 95–102. doi:10.1124/mol.106.033043. PMID 17440117.
- ↑ Hendricson AW, Maldve RE, Salinas AG, et al. (April 2007). "Aberrant synaptic activation of N-methyl-D-aspartate receptors underlies ethanol withdrawal hyperexcitability". J. Pharmacol. Exp. Ther. 321 (1): 60–72. doi:10.1124/jpet.106.111419. PMID 17229881.
- ↑ Dodd PR, Buckley ST, Eckert AL, Foley PF, Innes DJ (August 2006). "Genes and gene expression in the brains of human alcoholics". Ann. N. Y. Acad. Sci. 1074 (1): 104–15. doi:10.1196/annals.1369.010. PMID 17105908.
- ↑ Sircar R, Sircar D (May 2006). "Repeated ethanol treatment in adolescent rats alters cortical NMDA receptor". Alcohol. 39 (1): 51–8. doi:10.1016/j.alcohol.2006.07.002. PMID 16938629.
- ↑ Klein G, Gardiwal A, Schaefer A, Panning B, Breitmeier D (September 2007). "Effect of ethanol on cardiac single sodium channel gating". Forensic Sci. Int. 171 (2-3): 131–5. doi:10.1016/j.forsciint.2006.10.012. PMID 17129694.
- ↑ Shiraishi M, Harris RA (June 2004). "Effects of alcohols and anesthetics on recombinant voltage-gated Na+ channels". J. Pharmacol. Exp. Ther. 309 (3): 987–94. doi:10.1124/jpet.103.064063. PMID 14978193.
- ↑ Forrest MD (April 2015). "Simulation of alcohol action upon a detailed Purkinje neuron model and a simpler surrogate model that runs >400 times faster". BMC Neuroscience. 16 (27). doi:10.1186/s12868-015-0162-6.
- ↑ Forrest, Michael (April 2015). "The neuroscience reason we fall over when drunk". Science 2.0. Retrieved 24 June 2015.
- ↑ Farber, N. B.; Heinkel, C.; Dribben, W. H.; Nemmers, B.; Jiang, X. (2004). "In the adult CNS, ethanol prevents rather than produces NMDA antagonist-induced neurotoxicity". Brain Research. 1028 (1): 66–74. doi:10.1016/j.brainres.2004.08.065. PMID 15518643.
- ↑ DeGrandpre, Richard J.; Bickel, Warren K. (1996). Drug Policy and Human Nature: Psychological Perspectives on the Prevention, Management, and Treatment of Illicit Drug Abuse. New York: Plenum Press. ISBN 0-306-45241-3.
- ↑ Latest Buzz in Research: Intoxicated Honey bees may clue Scientists into Drunken Human Behavior, The Ohio State Research News, Research Communications, Columbus OH, 23 October 2004.
- ↑ Abramson CI, Stone SM, Ortez RA, et al. (August 2000). "The development of an ethanol model using social insects I: behavior studies of the honey bee (Apis mellifera L.)". Alcohol. Clin. Exp. Res. 24 (8): 1153–66. doi:10.1111/j.1530-0277.2000.tb02078.x. PMID 10968652.
- ↑ Intoxicated Honey Bees May Clue Scientists Into Drunken Human Behavior, Science Daily, 25 October 2004
- ↑ Abramson CI, Fellows GW, Browne BL, Lawson A, Ortiz RA (April 2003). "Development of an ethanol model using social insects: II. Effect of Antabuse on consumatory responses and learned behavior of the honey bee (Apis mellifera L.)". Psychol Rep. 92 (2): 365–78. doi:10.2466/PR0.92.2.365-378. PMID 12785614.
External links
- Global Status Report on Alcohol 2004 by the World Health Organization.
- Heberlein, Ulrike; Wolf, Fred W.; Rothenfluh, Adrian; Guarnieri, Douglas J. (2004-01-08). "Molecular Genetic Analysis of Ethanol Intoxication in Drosophila Melanogaster". Integrative and Comparative Biology. 44 (4): 269–74. doi:10.1093/icb/44.4.269. PMID 21676709.
- Jane behaving badly (New Scientist article on short-term effects of alcohol)