Sexual selection in mammals

Sexual selection in mammals started with Charles Darwin’s observations concerning sexual selection, including sexual selection in humans, and in other mammals,[1] consisting of male-male competition and mate choice that mold the development of future phenotypes in a population for a given species.[2][3]
Elephant seals

A good example of intrasexual selection, in which males fight for dominance over a harem of females, is the elephant seal - large, oceangoing mammals of the genus Mirounga. There are two species: the northern (M. angustirostris) and southern elephant seal (M. leonina) - the largest carnivore living today. Both species show extreme sexual dimorphism, possibly the largest of any mammal, with southern males typically five to six times heavier than the females.[4] While the females average 400 to 900 kilograms (880 to 1,980 lb) and 2.6 to 3 metres (8.5 to 9.8 ft) long, the bulls average 2,200 to 4,000 kilograms (4,900 to 8,800 lb) and 4.2 to 5 metres (14 to 16 ft) long.[5][6] The record-sized bull, shot in Possession Bay, South Georgia, on February 28, 1913, measured 6.85 metres (22.5 ft) long and was estimated to weigh 5,000 kilograms (11,000 lb).[7][8] The maximum weight of a female is 1,000 kilograms (2,200 lb) with a length of 3.7 metres (12 ft).
Males arrive in the colonies before the females and fight for control of harems.[9] Large body size confers advantages in fighting. The agonistic behaviour of the bulls gives rise to a dominance hierarchy, with access to harems and breeding activity being determined by rank.[10] The dominant bulls or "harem masters" establish harems of several dozen females. The least successful males have no harems, but may try to copulate with a harem male's females when the dominant male is not looking. A dominant male must stay in his territory to defend it, which can mean months without eating, living on his store of blubber. Some males have stayed ashore for more than three months without food. Two fighting males use their weight and canine teeth against each other. The outcome is rarely fatal, and the defeated bull will flee; however, bulls suffer severe tears and cuts. Males commonly vocalize with a coughing roar that serves in both individual recognition and size assessment. Conflicts between high-ranking males are more often resolved with posturing and vocalizing than with physical contact.[10]

In the case of intrasexual selection, adorned males may gain a reproductive advantage without the intervention of female preference. This advantage will be conferred by weapons used in the process of resolving disputes, such as those over territorial rights. The use of sexual ornamentation as a signaling device to create a dominance hierarchy among males, also known as a pecking order, allows struggle to proceed without excessive injury or fatality. It is predominantly when two opposing males are so closely matched, as would be found in males not having established themselves in a dominance hierarchy, that asymmetries cannot be found and the confrontation escalates to a point where the asymmetries must be proved by aggressive use of ornamentation.
How often males will physically engage each other, and in what manner, can best be understood by applying game theory developed for biology, most notably by John Maynard Smith.[11]
An uncertain example: the giraffe

The evolutionary origins of the giraffe's (Giraffa camelopardalis) long neck are controversial. The long-accepted "competing browser's hypothesis" originally put forth by Charles Darwin has been put into question. Originally, scientists believed that the elongation of the giraffe's neck had been a result of natural selection acting in relation to foraging behaviour, where it was supposed that longer necks enabled favoured individuals to gather food inaccessible to other animals.[12] But even though the giraffe’s overall height is about 6 meters, it still typically feeds at about 2 meters above the ground.[13] Moreover, the giraffe's kudu, impala, and steenbok competitors do not feed above 2 meters and prefer feeding at shoulder level as well, rather than at the maximum height they could reach.[14]
An alternative explanation for the origin of long necks in giraffe is sexual selection. Male giraffe often neck with other males to exhibit dominance.[15] There are six criteria that need to be satisfied for the exaggerated neck to be classified as a result of sexual selection. The characteristic should be more exaggerated in one of the sexes; it must be used to indicate dominance; have no direct survival benefits; cost the organism in terms of survival or other factors (e.g., energetics[16][17]); positive allometry should be observed.[18] But evolutionary history shows that increased neck length is not correlated to increases in other parts of the body, which would be expected from foraging selection, so sexual selection may be a more satisfactory explanation.[19] Studies have failed to resolve the causes involved: perhaps the neck was a result of both or other forces.[20]
Precopulatory mechanisms
Precopulatory mechanisms determine who father an offspring prior to sex. Male-male competition is the biggest precopulatory mechanism in mammals. Sexual dimorphism is a result of male-male competition that is easily seen in species.
Male-male competition

Male-male competition to copulate with the opposite sex is often seen in mammals. African elephants strongly promote male-male competition. Elephants continuously grow throughout their lifetime. As males grow older, they also experience increasing lapses of musth, a violent sexual excitement, and most reproductive success happens to males in musth as it helps them win fights.[21] A fight between a male in musth and one not can result in the death of the latter.
Species with intense male-male competition are known to exhibit the most size dimorphism.[22] For example, female American black bears (Ursus americanus) are 20-40% smaller than males.[23]
Male mammals can compete for harems as well with elephant seals competing fiercely for harems. As mammals reach sexual maturity, secondary sexual characteristic arise.[24] Elephant seals have a proboscis in the adult male, which is used to project loud noises, frequently heard during the mating season.[25] Elephant seals with a bigger proboscis emit lower sounds than males with a smaller proboscis and are the bigger of the males in a colony. Mate-guarding is an important factor in male-male competition to ensure fertilization of an offspring, and, when successful, helps to overlook and court the female.[26] It especially prevents sperm competition from occurring as it ensures reproductive success. This process can be engaged when a post-coital signal is sent to a male to keep guard by the female.[27] Mating plugs are a form of mate-guarding that have proved to show precopulatory female choice.[28] Copulatory plugs are commonly acellular and thought to be made by proteins from the seminal vesicles.[29] DNA taken from copulatory plugs show females deter from mating with close relatives.
Callings

During the breeding season, mammals will call out to the opposite sex. Male koalas that are bigger will let out a different sound than smaller koalas. The bigger males which are routinely sought out for are called sires. Females choose sires because of indirect benefits that their offspring could inherit, like larger bodies.[30] Non-sires and females do not vary in their body mass and can reject a male by screaming or hitting him. Male-male competition is rarely exhibited in koalas.[31] Acoustic signaling is a type of call that can be used from a significant distance encoding an organism’s location, condition and identity.[32] Sac-winged bats display acoustic signaling, which is often interpreted as songs. When females hear these songs, named a ‘whistle’, they call onto the males to breed with a screech of their own. This action is termed ‘calling of the sexes’.[33] Red deer and spotted hyenas along with other mammals also perform acoustic signaling.[34][35]
Testosterone

Testosterone is a driving factor towards achieving fertilizing success. Bighorn sheep display curved horns on the rams of the species which are big compared to the small horns displayed on the females of the species. The bigger the horns are, the more testosterone there was found to be in the male. This is important because social rank has a positive correlation with the length of the horns.[24] Social rank leads to tending of a group of females to copulate with.[36] Testosterone also appears more in polygynous species than in monogamous species.[37]
Polyandry
Polyandrous females have two or more mating partners while they are in heat. Females are more likely to find a new mate when their current male had a high number of paternities the year before or their current male was old. This is presumed to have an effect on offspring and giving them more genetic diversity.[38][39]
Sex-role reversal
_Tswalu.jpg)
Sex role reversal is the change in behavior of a specific species from their ordinary pattern. Sex-role reversal supports sexual dimorphism very strongly.[40] Female-female competition is a common abnormality within animals with accepted sex roles. Females invest into choosing the best possible mate because they have more of a part in bringing up their offspring than males (gestation and lactation). Gestation and lactation are energy-consuming, which means their competition for resources is high.[41] Female-female competitions are observed to gain access for better mates. Meerkat females acquire dominant status because resources for female reproduction are scarce.[42] Dominant females in this species are heavier and win in competitions over other females.
Postcopulatory mechanisms
Copulating with the opposite sex does not ensure fertilization of an egg. Postcopulatory mechanisms include sperm competition and cryptic female choice.
Sperm competition
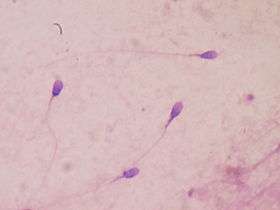
Sperm competition involves male gametes trying to fertilize eggs first. As a result of sperm competition, some males in a given species can develop bigger testes and seminal vesicles.[43] Larger midpiece areas in the sperm that contain mitochondria are also observed.[37] Larger testes and bigger midpieces in sperm are seen in females that mate with multiple partners. A female that has been with multiple partners will most likely give birth to an offspring fathered by the male that produced the most or faster sperm.[43] It was found that primates and rodents with longer flagellum fathered more offspring.[44]
Cryptic female choice
Cryptic female choice is a postcopulatory mechanism that cannot be observed because it takes place inside a female’s body.[45] It enables a female to have some control over who fathers her child even after fertilization.[46]
In some species, females may choose to mate with more than one male to prevent infanticide or harassment. Infanticide can be prevented by confusing the males in a given colony. If the female mates with multiple males, then the males will not know for sure who fathered the offspring. Infanticide can also be prevented by choosing a male that will protect her and the offspring.[43] Sexual harassment may be avoided if females give in to males and copulate when they please.[47]
References
- ↑ Jones, A. G. & Ratterman, N. L. Mate choice and sexual selection: what have we learned since Darwin? Proceedings of the National Academy of Sciences of the United States of America 106 Suppl , 10001–8 (2009)
- ↑ Cyrus Chu, C. Y. & Lee, R. D. Sexual dimorphism and sexual selection: a unified economic analysis. Theoretical population biology 82, 355–63 (2012)
- ↑ Vogt, Yngve (January 29, 2014). "Large testicles are linked to infidelity". Phys.org. Retrieved January 31, 2014.
- ↑ Perrin, William F.; Würsig, Bernd; Thewissen, J. G. M., eds. (2008-11-24). "Earless Seals". Encyclopedia of Marine Mammals (2nd ed.). Burlington, Massachusetts: Academic Press. p. 346. ISBN 978-0-12-373553-9.
- ↑ "Southern Elephant Seal". pinnipeds.org. Seal Conservation Society.
- ↑ Block, D.; Meyer, Philip; Myers, P. (2004). "Mirounga leonina (Southern elephant seal)". Animal Diversity Web. The Regents of the University of Michigan. Retrieved September 11, 2010.
- ↑ Wood, The Guinness Book of Animal Facts and Feats. Sterling Pub Co Inc (1983), ISBN 978-0-85112-235-9
- ↑ Carwardine, Mark (2008). Animal Records. New York: Sterling. p. 61. ISBN 978-1-4027-5623-8.
- ↑ Jones, E. (1981). "Age in relation to Breeding Status of the Male Southern Elephant Seal, Mirounga leonina (L.), at Macquarie Island". Australian Wildlife Research. 8 (2): 327–334. doi:10.1071/wr9810327.
- 1 2 McCann TS (1981). "Aggression and sexual activity of male Southern elephant seals, Mirounga leonina". Journal of Zoology. 195 (3): 295–310. doi:10.1111/j.1469-7998.1981.tb03467.x.
- ↑ Maynard Smith, J (1982) Evolution and the Theory of Games. Cambridge University Press, pp. 131-137. ISBN 0-521-28884-3
- ↑ Mitchell, G.; Skinner, J. D. (200). "On the origin, evolution and phylogeny of giraffes Giraffa camelopardalis". Transactions of the Royal Society of South Africa. 58: 51–73. doi:10.1080/00359190309519935.
- ↑ Leuthold, B. M.; Leuthold, W. (1972). "Food habits of giraffe in Tsavo National Park, Kenya". African Journal of Ecology. 10 (2): 129–141. doi:10.1111/j.1365-2028.1972.tb00867.x.
- ↑ Simmons, R. E.; Altwegg, R. (2010). "Necks-for-sex or competing browsers? A critique of ideas on the evolution of giraffe". Journal of Zoology. 282: 6–12. doi:10.1111/j.1469-7998.2010.00711.x.
- ↑ Mitchell, G.; Skinner, J. D.; S. J. van Sittert (2009). "Sexual selection is not the origin of long necks in giraffes". Journal of Zoology. 278 (4): 281–286. doi:10.1111/j.1469-7998.2009.00573.x.
- ↑ Oufiero, C. E., and T. Garland. 2007. Evaluating performance costs of sexually selected traits. Functional Ecology 21:676-689.
- ↑ Husak, J. F., and J. G. Swallow. 2011. Compensatory traits and the evolution of male ornaments. Behaviour 148:1–29.
- ↑ Simmons, R. E.; Scheepers, L. (1996). "Winning by a neck: Sexual selection in the evolution of giraffe". American Naturalist. 148 (5): 771–786. doi:10.1086/285955.
- ↑ Senter, P. (2007). "Necks for sex: sexual selection as an explanation for sauropod dinosaur neck elongation". Journal of Zoology. 271: 45–53. doi:10.1111/j.1469-7998.2006.00197.x.
- ↑ Langman, V. A. (1977). "Cow-calf relationships in giraffe (Giraffa-Camelopardalis-Giraffa)". Zeitschrift für Tierpsychologie. 43: 264–286.
- ↑ Hollister-Smith, J. a. et al. Age, musth and paternity success in wild male African elephants, Loxodonta africana. Animal Behaviour 74, 287–296 (2007)
- ↑ Floyd, W. Sexual-size dimorphism : Influence of mass and mating systems in the most dimorphic mamma. (1998)
- ↑ Kovach, A. I. & Powell, R. A. Effects of body size on male mating tactics and paternity in black bears , Ursus americanus. 1268, 1257–1268 (2003)
- 1 2 Martin, A. M., Presseault-Gauvin, H., Festa-Bianchet, M. & Pelletier, F. Male mating competitiveness and age-dependent relationship between testosterone and social rank in bighorn sheep. Behavioral Ecology and Sociobiology (2013).doi:10.1007/s00265-013-1516-7
- ↑ Sanvito, S., Galimberti, F. & Miller, E. H. Having a big nose : structure , ontogeny , and function of the elephant seal proboscis. 220, 207–220 (2007)
- ↑ Schubert, M., Schradin, C., Rödel, H. G., Pillay, N. & Ribble, D. O. Male mate guarding in a socially monogamous mammal, the round-eared sengi: on costs and trade-offs. Behavioral Ecology and Sociobiology 64, 257–264 (2009)
- ↑ Maestripieri, D., Leoni, M., Raza, S. S., Hirsch, E. J. & Whitham, J. C. Female Copulation Calls in Guinea Baboons: Evidence for Postcopulatory Female Choice? International Journal of Primatology 26, 737–758 (2005)
- ↑ All, A. M. & Terms, J. Sperm Competition and the Evolution of Nonfertilizing Sperm in Mammals Author ( s ): A . H . Harcourt Published by : Society for the Study of Evolution Stable URL : http://www.jstor.org/stable/2409666 . AND THE EVOLUTION SPERM IN MAMMALS. 45, 314–328 (2013)
- ↑ McCreight, J. C., DeWoody, J. a. & Waser, P. M. DNA from copulatory plugs can give insights into sexual selection. Journal of Zoology 284, 300–304 (2011)
- ↑ Charlton, B. D., Ellis, W. a. H., Brumm, J., Nilsson, K. & Fitch, W. T. Female koalas prefer bellows in which lower formants indicate larger males. Animal Behaviour 84, 1565–1571 (2012)
- ↑ Ellis, W. a. H. & Bercovitch, F. B. Body size and sexual selection in the koala. Behavioral Ecology and Sociobiology 65, 1229–1235 (2011)
- ↑ Wilkins, M. R., Seddon, N. & Safran, R. J. Evolutionary divergence in acoustic signals: causes and consequences. Trends in ecology & evolution 28, 156–66 (2013)
- ↑ Ecology, E. SONGS , SCENTS , AND SENSES : SEXUAL SELECTION IN THE GREATER SAC-WINGED BAT , SACCOPTERYX BILINEATA. 89, 1401–1410 (2008)
- ↑ Logan, C. J. & Clutton-Brock, T. H. Validating methods for estimating endocranial volume in individual red deer (Cervus elaphus). Behavioural processes 92, 143–6 (2013)
- ↑ Goller, K. V, Fickel, J., Hofer, H., Beier, S. & East, M. L. Coronavirus genotype diversity and prevalence of infection in wild carnivores in the Serengeti National Park, Tanzania. Archives of virology 158, 729–34 (2013)
- ↑ Mating in Bighorn Sheep : Multiple Creative Male Strategies Author ( s ): John T . Hogg Published by : American Association for the Advancement of Science Stable URL : http://www.jstor.org/stable/1694008 . 225, 526–529 (2013)
- 1 2 Dixson, A. F. & Anderson, M. J. Sexual behavior, reproductive physiology and sperm competition in male mammals. Physiology & behavior 83, 361–71 (2004)
- ↑ Kvarnemo, C., Simmons, L. W. & B, P. T. R. S. Polyandry as a mediator of sexual selection before and after mating Polyandry as a mediator of sexual selection before and after mating. (2013)
- ↑ Briefer, E. F., Farrell, M. E., Hayden, T. J. & McElligott, A. G. Fallow deer polyandry is related to fertilization insurance. Behavioral Ecology and Sociobiology 67, 657–665 (2013)
- ↑ Ah-King, M. & Ahnesjö, I. The “Sex Role” Concept: An Overview and Evaluation. Evolutionary Biology (2013).doi:10.1007/s11692-013-9226-7
- ↑ Rosvall, K. a Intrasexual competition in females: evidence for sexual selection? Behavioral ecology : official journal of the International Society for Behavioral Ecology 22, 1131–1140 (2011)
- ↑ Clutton-Brock, T. H. et al. Intrasexual competition and sexual selection in cooperative mammals. Nature 444, 1065–8 (2006)
- 1 2 3 Plavcan, J. Sexual dimorphism in primate evolution. American journal of physical anthropology 53, 25–53 (2001)
- ↑ Gomendio, M. & Roldan, E. R. Sperm competition influences sperm size in mammals. Proceedings. Biological sciences / The Royal Society 243, 181–5 (1991)
- ↑ All, P. M. & Terms, J. Cryptic Female Choice : Criteria for Establishing Female Sperm Choice Author ( s ): T . R . Birkhead Published by : Society for the Study of Evolution Stable URL : http://www.jstor.org/stable/2411251 . CRYPTIC FEMALE CHOICE : CRITERIA FOR ESTABLISHING FEM. 52, 1212–1218 (2013)
- ↑ Holt, W. V & Fazeli, A. The oviduct as a complex mediator of mammalian sperm function and selection. Molecular reproduction and development 77, 934–43 (2010)
- ↑ Wolff, J. O. & Macdonald, D. W. Promiscuous females protect their offspring. Trends in ecology & evolution 19, 127–34 (2004)