Plasmodium
| Plasmodium | |
|---|---|
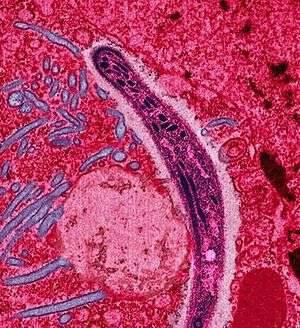 | |
| False-colored electron micrograph of a Plasmodium sp. sporozoite. | |
| Scientific classification | |
| (unranked): | SAR |
| (unranked): | Alveolata |
| Phylum: | Apicomplexa |
| Class: | Aconoidasida |
| Order: | Haemosporida |
| Family: | Plasmodiidae |
| Genus: | Plasmodium Marchiafava & Celli, 1885 |
| Species | |
Plasmodium is a genus of parasitic protozoa, many of which cause malaria in their hosts.[1] The parasite always has two hosts in its life cycle: a Dipteran insect host and a vertebrate host. Sexual reproduction always occurs in the insect, making it the definitive host.[2]
The life-cycles of Plasmodium species involve several different stages both in the insect and the vertebrate host. These stages include sporozoites, which are injected by the insect vector into the vertebrate host's blood. Sporozoites infect the host liver, giving rise to merozoites and (in some species) hypnozoites. These move into the blood where they infect red blood cells. In the red blood cells, the parasites can either form more merozoites to infect more red blood cells, or produce gametocytes which are taken up by insects which feed on the vertebrate host. In the insect host, gametocytes merge to sexually reproduce. After sexual reproduction, parasites grow into new sporozoites, which move to the insect's salivary glands, from which they can infect a vertebrate host bitten by the insect.[1]
The genus Plasmodium was first described in 1885. It now contains about 200 species, which are spread across the world where both the insect and vertebrate hosts are present. Five species regularly infect humans, while many others infect birds, reptiles, rodents, and various primates.
History
Plasmodia were first identified when Charles Louis Alphonse Laveran described parasites in the blood of malaria patients in 1880.[3] He named the parasite Oscillaria malariae.[3] The fact that several species may be involved in causing different forms of malaria was first recognized by Camillo Golgi in 1886.[3] Soon thereafter, Giovanni Batista Grassi and Raimondo Filetti named the parasites causing two different types of human malaria Plasmodium vivax and Plasmodium malariae.[3] In 1897, William Welch identified and named Plasmodium falciparum. This was followed by the recognition of the other two species of Plasmodium which infect humans: Plasmodium ovale (1922) and Plasmodium knowlesi (identified in long-tailed macaques in 1931; in humans in 1965).[3] The contribution of insect hosts to the Plasmodium life cycle was described in 1897 by Ronald Ross and in 1899 by Giovanni Batista Grassi, Amico Bignami and Giuseppe Bastianelli.[3]
Life cycle
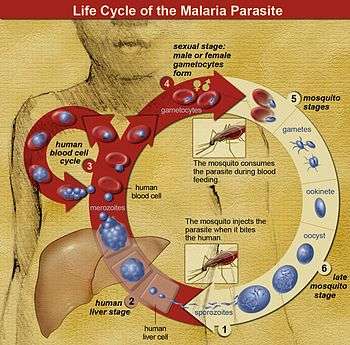
The life cycle of Plasmodium involves several distinct stages in the insect and vertebrate hosts. In infected mosquitoes, parasites in the salivary gland are called sporozoites. When the mosquito bites a vertebrate host, sporozoites are injected into the host with the saliva. From there, the sporozoites enter the bloodstream and are transported to the liver, where they invade and replicate within hepatocytes. At this point, some species of Plasmodium can form a long-lived dormant stage called a hypnozoite which can remain in the liver for many years.[2] The parasites that emerge from infected hepatocytes are called merozoites, and these return to the blood to infect red blood cells.
Within the red blood cells, the merozoites grow first to a ring-shaped form and then to a larger form called a trophozoite. Trophozoites then mature to schizonts which divide several times to produce new merozoites. The infected red blood cell eventually bursts, allowing the new merozoites to travel within the bloodstream to infect new red blood cells. Most merozoites continue this replicative cycle, however some merozoites differentiate into male or female sexual forms called gametocytes. These gametocytes circulate in the blood until they are taken up when a mosquito feeds on the infected vertebrate host, taking up blood which includes the gametocytes.[2]
In the mosquito, the gametocytes move along with the blood meal to the mosquito's midgut. Here the gametocytes develop into male and female gametes which fertilize each other, forming a zygote. Zygotes then develop into a motile form called an ookinete, which penetrates the wall of the midgut. Upon traversing the midgut wall, the ookinete embeds into the gut's exterior membrane and develops into an oocyst. Oocysts divide many times to produce large numbers of small elongated sporozoites. These sporozoites migrate to the salivary glands of the mosquito where they can be injected into the blood of the next host the mosquito bites, repeating the cycle.[2]
Description

Plasmodium species each have 14 chromosomes in the nucleus, as well as genetic material in the mitochondrion and in the apicoplast. The chromosomes vary from 500 kilobases to 3.5 megabases in length. The apicoplast is involved in isoprenoid metabolism, Fe-S cluster synthesis, fatty acid synthesis, and phospholipid biosynthesis.[4]
On a molecular level, the parasite damages red blood cells using plasmepsin enzymes — aspartic acid proteases which degrade hemoglobin.
Taxonomy
Plasmodium is a member of the family Plasmodiidae, order Haemosporidia and phylum Apicomplexa which, along with dinoflagellates and ciliates, make up the taxonomic group Alveolata.[5]
Plasmodium species were originally classified into subgenera based on their morphology, location, and host specificity. However, more recent studies of Plasmodium species using molecular methods have occasionally given results which conflict with the original taxonomic system.[6]
Evolution
Evolutionary relationships of species within the genus Plasmodium have been controversial.[6] Plasmodium species were originally divided by morphology, life-cycle characteristics, and host species. However, modern molecular approaches for determining evolutionary relationships have given results which conflict with older classification methods.[6] Many attempts to clarify Plasmodium taxonomy with molecular methods have also run into technical challenges. Ribosomal RNA sequencing, which is often used in other organisms to determine evolutionary relationships, is challenging to interpret from Plasmodium species as Plasmodia maintain several different copies of ribosomal RNA which are expressed at different stages of the life cycle and which may be able to recombine with one another.[6] Another commonly used marker for evolutionary studies has been the circumsporozoite protein (CSP) which is present in all Plasmodium species.[6] However, analyses of CSP sequences are complicated by the fact that the sequence of CSP, which is present on the surface of the parasite during infection, is under heavy selective pressure from the host immune system.[6] This could potentially obscure relevant changes.[6]
Environmental factors play a considerable role in the evolution of Plasmodium and the transmission of malaria. The genetic information of Plasmodium falciparum has signaled a recent expansion that coincides with the agricultural revolution. It is likely that the development of extensive agriculture increased mosquito population densities by giving rise to more breeding sites, which may have triggered the evolution and expansion of Plasmodium falciparum.[7]
There are over one hundred species of mosquito-transmitted Plasmodium. The phylogeny of these malarial parasites suggests that the Plasmodium of mammalian hosts forms a well-defined clade strongly associated with the specialization to the Anopheles mosquito vector. This was a major evolutionary transition that allowed Plasmodium to exploit humans and other mammals.[8][9]
P. falciparum, the most lethal malaria parasite of humans, evolved from a "nearly identical" parasite of western gorillas.[10]
The high mortality and morbidity caused by malaria—especially that caused by P. falciparum—has placed the greatest selective pressure on the human genome in recent history. Several genetic factors provide some resistance to Plasmodium infection, including sickle cell trait, thalassaemia traits, glucose-6-phosphate dehydrogenase deficiency, and the absence of Duffy antigens on red blood cells.[11] [12]
Although there are therapeutic medications to treat malaria, Plasmodium has accumulated increasing drug resistance over time. A recent examination has shown that even artemisinin, one of the most powerful anti-malarial drugs, has been experiencing decreased efficacy due to the development of resistance.[13]
Subgenera
Plasmodium species have been subdivided into subgenera, which group species of similar morphology and with similar hosts.
- Subgenus Asiamoeba (lizards)
- Subgenus Bennettinia (birds)
- Subgenus Carinamoeba (reptiles)
- Subgenus Giovannolaia (birds)
- Subgenus Haemamoeba (birds)
- Subgenus Huffia (birds)
- Subgenus Lacertamoeba (reptiles)
- Subgenus Laverania (higher primates)
- Subgenus Novyella (birds)
- Subgenus Paraplasmodium (lizards)
- Subgenus Plasmodium (monkeys, higher primates)
- Subgenus Sauramoeba (reptiles)
- Subgenus Vinckeia (non-primate mammals)
Species infecting monkeys and apes (the higher primates) with the exceptions of P. falciparum and P. reichenowi (which together make up the subgenus Laverania) are classified in the subgenus Plasmodium. Parasites infecting other mammals including lower primates (lemurs and others) are classified in the subgenus Vinckeia. The distinction between P. falciparum and P. reichenowi and the other species infecting higher primates was based on morphological findings but have since been confirmed by DNA analysis.
The four subgenera Giovannolaia, Haemamoeba, Huffia and Novyella were created by Corradetti et al.[14] for the known avian malarial species. A fifth — Bennettinia — was created in 1997 by Valkiunas.[15] The relationships between the subgenera are a matter of current investigation.[16]
P. juxtanucleare is the only member of the subgenus Bennettinia.
Unlike the mammalian and bird malarias those affecting reptiles have been more difficult to classify. In 1966 Garnham classified those with large schizonts as Sauramoeba, those with small schizonts as Carinamoeba and the single then-known species infecting snakes (Plasmodium wenyoni) as Ophidiella.[17] In 1988, Telford used this scheme as the basis for the current system.[18]
Hosts
All Plasmodia are parasitic and require both a vertebrate host and an insect host to reproduce. Known vertebrate hosts include various primates (including humans), birds, rodents, bats, porcupines and squirrels. Mosquitoes of the genera Culex, Anopheles, Culiseta, Mansonia and Aedes often serve as insect hosts for various Plasmodium species.
Humans

The species of Plasmodium that infect humans include:
- Plasmodium falciparum (the cause of malignant tertian malaria)
- Plasmodium vivax (the most frequent cause of benign tertian malaria)
- Plasmodium ovale (the other, less frequent, cause of benign tertian malaria)
- Plasmodium malariae (the cause of benign quartan malaria)
- Plasmodium knowlesi (the cause of severe quotidian malaria in South East Asia since 1965)
P. falciparum, P. vivax, P. ovale, and P. malariae together account for nearly all human infections with Plasmodium species, with P. falciparum accounting for the overwhelming majority of malaria deaths. An increasing number of cases of severe malaria in Southeast Asia have been attributed to P. knowlesi.
With the use of the polymerase chain reaction additional species have been identified in humans, although whether these species can regularly infect humans is not known. An experimental infection of a human volunteer with Plasmodium eylesi and Plasmodium cynomolgi has been reported, although the infection was not able to be transferred to another susceptible host, suggesting the parasites may not be able to productively infect humans.[19] A possible infection with Plasmodium tenue has also been reported, however doubts have been raised as to the validity of this diagnosis.[20] Other species which have been reportedly isolated from humans include Plasmodium brasilianum, Plasmodium inui, Plasmodium rhodiani, Plasmodium schweitzi, Plasmodium semiovale, and Plasmodium simium.
In addition to humans, P. vivax can infect chimpanzees and orangutans. In these hosts, infection tends to not cause severe disease, but may persist for some time.[21]
P. ovale can be transmitted to chimpanzees, and is found in Africa, the Philippines and New Guinea.
Non-human primates

The species that infect primates other than humans include: P. bouillize, P. brasilianum, P. bucki, P. cercopitheci,P. coatneyi, P. coulangesi, P. cynomolgi, P. eylesi, P. fieldi, P. foleyi, P. fragile, P. girardi, P. georgesi, P. gonderi, P. hylobati, P. inui, P. jefferyi, P. joyeuxi, P. knowlesi, P. lemuris, P. percygarnhami, P. petersi, P. reichenowi, P. rodhaini, P. sandoshami, P. semnopitheci, P. silvaticum, P. simiovale, P. simium, P. uilenbergi, P. vivax and P. youngei.
Many Plasmodium species infect more than one primate host species. Primates which have been found to be infected with Plasmodium include species of the genera Alouatta (also known as howler monkeys), Ateles (spider monkeys), Brachyteles (muriqui or wooly spider monkeys), Callicebus (titi monkeys), Chiropotes (bearded sakis), Lagothrix (woolly monkeys), Lemur (lemurs), Macaca (macaques), Pan (chimpanzees), Pongo (orangutans), and Saimiri (squirrel monkeys).[22][23]
The insect hosts of the Plasmodium species which infect primates are various species of Anopheles mosquitoes. Different species of Plasmdodium generally infect different species of mosquito, although some mosquito species can carry several Plasmodium species.
Non-primate mammals

The subgenus Vinckeia was created by Cyril Garnham to accommodate the mammalian parasites other than those infecting primates. Species infecting lemurs have also been included in this subgenus. Plasmodium species can infect a wide variety of mammals including rodents, ungulates, and bats. Several of the species which infect rodents have been shown to also infect the mosquito Anopheles stephensi.
P. aegyptensis, P. bergei, P. chabaudi, P. inopinatum, P. yoelli and P. vinckei infect rodents. P. bergei, P. chabaudi, P. yoelli and P. vinckei have been used to study malarial infections in the laboratory. Other members of this subgenus infect other mammalian hosts.
Birds

Species in five Plasmodium subgenera infect birds — Bennettinia, Giovannolaia, Haemamoeba, Huffia and Novyella.[24] Giovannolaia appears to be a polyphyletic group and may be sudivided in the future.[16] Species that infect birds include P. accipiteris, P. alloelongatum, P. anasum, P. ashfordi, P. bambusicolai, P. bigueti, P. biziurae, P. buteonis, P. cathemerium, P. circumflexum, P. coggeshalli, P. corradettii, P. coturnix, P. dissanaikei, P. durae, P. elongatum, P. fallax, P forresteri, P. gallinacium, P. garnhami, P. giovannolai, P. griffithsi, P. gundersi, P. guangdong, P. hegneri, P. hermani, P. hexamerium, P. huffi, P. jiangi, P. juxtanucleare, P. kempi, P. lophurae, P.lutzi, P. matutinum, P. nucleophilum, P. papernai, P. paranucleophilum, P. parvulum, P. pediocetti, P. paddae, P. pinotti, P. polare, P. relictum, P. rouxi, P. tenue, P. tejerai, P. tumbayaensis and P. vaughani.[25]
These infect a variety of bird species. In general each species of Plasmodium infects one to a few species of birds. Each species is also generally transmitted by a single insect species. Insect hosts include Aedes aegypti, Mansionia crassipes, and various Culex species.
Reptiles

Species in the subgenera Asiamoeba, Carinamoeba, Lacertaemoba, Paraplasmodium and Sauramoeba infect reptiles.[26] These species of Plasmodium include: P. achiotense, P. aeuminatum, P. agamae, P. arachniformis, P. attenuatum,P. aurulentum, P. australis, P. azurophilum, P. balli, P. basilisci, P. beebei, P. beltrani , P. brumpti, P. brygooi, P. chiricahuae, P. circularis, P. cnemaspi, P. cnemidophori, P. colombiense, P. cordyli, P. diminutivum, P. diploglossi, P. egerniae, P. fairchildi, P. floridense, P. gabaldoni, P. giganteum, P. gologoense, P. gracilis, P. guyannense, P. heischi, P. holaspi, P. icipeensis, P. iguanae, P. josephinae, P. kentropyxi, P. lacertiliae, P. lainsoni, P. lepidoptiformis, P. lionatum, P. loveridgei, P. lygosomae, P. mabuiae, P. mackerrasae, P. maculilabre, P. marginatum, P. mexicanum, P. michikoa, P. minasense, P. pelaezi, P. pessoai, P. pifanoi, P. pitmani, P. rhadinurum, P. sasai,P. saurocaudatum, P. scorzai, P. siamense, P. robinsoni, P. sasai, P. scorzai, P. tanzaniae, P. tomodoni, P. torrealbai, P. tribolonoti, P. tropiduri, P. uluguruense, P. uzungwiense, P. vacuolatum, P. vastator, P. volans, P. wenyoni and P. zonuriae.
Plasmodium species have been reported from over 3200 species of lizard and 29 species of snake. Only three species — P. pessoai, P. tomodoni and P. wenyoni — infect snakes. Species infecting lizards have been reported in relatively few insect hosts, including Lutzomyia and Culicoides species, Culex fatigans and Aedes aegypti.
Insects
Mosquitoes of the genera Culex, Anopheles, Culiseta, Mansonia and Aedes may act as insect hosts for various Plasmodia species. The best studies of these have been the Anopheles mosquitoes which host Plasmodia which cause human malaria, as well as Culex mosquitoes which host the Plasmodia that cause malaria in birds. In all cases, only female mosquitoes can be infected with Plasmodia species, since only the females feed on the blood of vertebrate hosts.
The survivorship and relative fitness of mosquitoes are not adversely affected by Plasmodium infection, which indicates the importance of vector fitness in shaping the evolution of Plasmodium.[27] Plasmodium has evolved the capability to manipulate mosquito feeding behavior. Mosquitoes harboring Plasmodium have a higher propensity to bite than uninfected mosquitoes. This tendency has facilitated the spread of Plasmodium to the various hosts.[28]
Species reclassified into other genera
Several species of Plasmodium have been reclassified, mostly to Hepatocystis. These include:
- Plasmodium epomophori to Hepatocystis epomophori
- Plasmodium kochi to Hepatocystis kochi
- Plasmodium limnotragi to Hepatocystis limnotragi (Van Denberghe 1937)
- Plasmodium pteropi to Hepatocystis pteropi (Breinl 1911)
- Plasmodium ratufae to Hepatocystis ratufae (Donavan 1920)
- Plasmodium vassali to Hepatocystis vassali (Laveran 1905)
- Plasmodium gonatodi to Garnia gonatodi
Link to Ebola virus
In August 2016, the National Institute Of Allergy and Infectious Diseases (NIAID) came out with a study that showed that people with the Ebola virus were 20 percent more likely to survive it if they were infected with the Plasmodium parasite. The study was published in Clinical Infectious Diseases. Researchers did their work at an ebola treatment unit in Monrovia, Liberia.[29] Researchers also found that the greater the number of Plasmodium parasites in the body, the greater the rate of Ebola survival. NIAID is part of the National Institutes of Health.[30]
Nomenclature
As with many other genera, the genus name also yields a common noun; thus species of the genus are known as plasmodia.
See also
References
- 1 2 "CDC - Malaria Parasites - About". CDC: Malaria. U.S. Centers for Disease Control and Prevention. Retrieved 28 December 2015.
- 1 2 3 4 "CDC - Malaria Parasites - Biology". CDC: Malaria. U.S. Centers for Disease Control and Prevention. Retrieved 28 December 2015.
- 1 2 3 4 5 6 "The History of Malaria, an Ancient Disease". U.S. Centers for Disease Control and Prevention. Retrieved 31 May 2016.
- ↑ vanDooren, G; Striepen, B (2013). "The Algal Past and Parasite Present of Apicoplast". Annual Review of Microbiology. 67: 271–289. doi:10.1146/annurev-micro-092412-155741. PMID 23808340.
- ↑ Morrison, David A. (2009). "Evolution of the Apicomplexa: Where are we now?". Trends in Parasitology. 25 (8): 375–82. doi:10.1016/j.pt.2009.05.010. PMID 19635681.
- 1 2 3 4 5 6 7 Zilversmit M; Perkins S. "Plasmodium". Tree of Life Web Project. Retrieved 1 June 2016.
- ↑ Hume, J.C.; Lyons, E.J.; Day, K.P. (2003). "Human migration, mosquitoes and the evolution of Plasmodium falciparum". Trends Parasitol. 19 (3): 144–9. doi:10.1016/s1471-4922(03)00008-4. PMID 12643998.
- ↑ Martinsen ES, Perkins SL, Schall JJ (2008). "A three-genome phylogeny of malaria parasites (Plasmodium and closely related genera): evolution of life-history traits and host switches". Molecular Phylogenetics and Evolution. 47 (1): 261–273. doi:10.1016/j.ympev.2007.11.012. PMID 18248741.
- ↑ Yotoko, K.S.C.; Elisei C. (November 2006). "Malaria parasites (Apicomplexa, Haematozoea) and their relationships with their hosts: is there an evolutionary cost for the specialization?". J. Zoo. Syst. Evol. Res. 44 (4): 265–273. doi:10.1111/j.1439-0469.2006.00377.x.
- ↑ Weimin Liu, Yingying Li, Gerald H. Learn, Rebecca S. Rudicell, Joel D. Robertson, Brandon F. Keele, Jean-Bosco N. Ndjango, Crickette M. Sanz, David B. Morgan, Sabrina Locatelli, Mary K. Gonder, Philip J. Kranzusch, Peter D. Walsh, Eric Delaporte, Eitel Mpoudi-Ngole, Alexander V. Georgiev, Martin N. Muller, George M. Shaw, Martine Peeters, Paul M. Sharp, Julian C. Rayner & Beatrice H. Hahn (23 September 2010). "Origin of the human malaria parasite Plasmodium falciparum in gorillas". Nature. 467 (7314): 420–425. doi:10.1038/nature09442. PMC 2997044
 . PMID 20864995.
. PMID 20864995. - ↑ Kwiatkowski DP (2005). "How malaria has affected the human genome and what human genetics can teach us about malaria". American Journal of Human Genetics. 77 (2): 171–92. doi:10.1086/432519. PMC 1224522. PMID 16001361.

- ↑ Hedrick PW (2011). "Population genetics of malaria resistance in humans". Heredity. 107 (4): 283–304. doi:10.1038/hdy.2011.16. PMC 3182497. PMID 21427751.
- ↑ Ashley, E.A.; Dhorda, M.; Fairhurst, R.M.; et al. (2014). "Spread of artemisinin resistance in Plasmodium falciparum malaria". N Engl J Med. 371 (5): 411–23. doi:10.1056/nejmoa1314981. PMC 4143591. PMID 25075834.
- ↑ Corradetti A.; Garnham P.C.C.; Laird M. (1963). "New classification of the avian malaria parasites". Parassitologia. 5: 1–4.
- ↑ Valkiunas G (1997). "Bird Haemosporidia". Acta Zoologica Lituanica. 3–5: 1–607. ISSN 1392-1657.
- 1 2 Martinsen ES, Waite JL, Schall JJ (April 2007). "Morphologically defined subgenera of Plasmodium from avian hosts: test of monophyly by phylogenetic analysis of two mitochondrial genes". Parasitology. 134 (Pt 4): 483–90. doi:10.1017/S0031182006001922. PMID 17147839.
- ↑ Garnham 1966
- ↑ Telford S (1988). "A contribution to the systematics of the reptilian malaria parasites, family Plasmodiidae (Apicomplexa: Haemosporina)". Bulletin of the Florida State Museum Biological Sciences. 34 (2): 65–96.
- ↑ Tsukamoto M (1977). "An imported human malarial case characterized by severe multiple infections of the red blood cells". Ann. Trop. Med. Parasitol. 19 (2): 95–104.
- ↑ Russel P.F. (1928). "Plasmodium tenue (Stephens): A review of the literature and a case report". Am. J. Trop. Med. s1–8 (5): 449–479.
- ↑ Reid MJ, Ursic R, Cooper D, et al. (December 2006). "Transmission of human and macaque Plasmodium spp. to ex-captive orangutans in Kalimantan, Indonesia". Emerging Infect. Dis. 12 (12): 1902–8. doi:10.3201/eid1212.060191. PMC 3291341. PMID 17326942.
- ↑ Collins WE, Sullivan JS, Nace D, Williams T, Williams A, Barnwell JW (February 2008). "Observations on the sporozoite transmission of Plasmodium vivax to monkeys". J. Parasitol. 94 (1): 287–8. doi:10.1645/GE-1283.1. PMID 18372652.
- ↑ Collins WE, Richardson BB, Morris CL, Sullivan JS, Galland GG (July 1998). "Salvador II strain of Plasmodium vivax in Aotus monkeys and mosquitoes for transmission-blocking vaccine trials". Am. J. Trop. Med. Hyg. 59 (1): 29–34. PMID 9684622.
- ↑ Wiersch SC, Maier WA, Kampen H (May 2005). "Plasmodium (Haemamoeba) cathemerium gene sequences for phylogenetic analysis of malaria parasites". Parasitol. Res. 96 (2): 90–4. doi:10.1007/s00436-005-1324-8. PMID 15812672.
- ↑ Clark, Nicholas; Clegg, Sonya; Lima, Marcos (2014). "A review of global diversity in avian haemosporidians (Plasmodium and Haemoproteus: Haemosporida): new insights from molecular data". International Journal for Parasitology. 44 (5): 329–338. doi:10.1016/j.ijpara.2014.01.004. PMID 24556563.
- ↑ Schall JJ (December 2000). "Transmission success of the malaria parasite Plasmodium mexicanum into its vector: role of gametocyte density and sex ratio". Parasitology. 121 (Pt 6): 575–80. doi:10.1017/s0031182000006818. PMID 11155927.
- ↑ Ferguson, HM; Mackinnon, MJ; Chan, BH; Read, AF. (2003). "Mosquito mortality and the evolution of malaria virulence". Evolution. 57 (12): 2792–804. doi:10.1554/03-211. PMID 14761058.
- ↑ Koella, J.C.; Sørensen, F.L.; Anderson, R.A. (1998). "The malaria parasite, Plasmodium falciparum, increases the frequency of multiple feeding of its mosquito vector, Anopheles gambiae". Proc Biol Sci. 265 (1398): 763–8. doi:10.1098/rspb.1998.0358. PMC 1689045. PMID 9628035.
- ↑ Murtha, Alex (2016-08-19). "NIAID researchers explore link between Ebola virus survival and malaria-causing parasite". Homeland Preparedness News. Retrieved 2016-08-29.
- ↑ "Media Availability: NIH Explores Connection Between Ebola Survival and Co-Infection with Malaria Parasites". www.niaid.nih.gov. 2016-08-16. Retrieved 2016-08-29.
Further reading
Identification
- Garnham, P.C.C. (1966). Malaria Parasites And Other Haemosporidia. Oxford: Blackwell. ISBN 0397601328.
- Hewitt, R.I. (1940). Bird Malaria. American Journal of Hygiene. 15. Baltimore: Johns Hopkins Press.
- Laird, M. (1998). Avian Malaria in the Asian Tropical Subregion. Singapore: Springer. ISBN 9813083190.
Biology
- Baldacci P, Ménard R (October 2004). "The elusive malaria sporozoite in the mammalian host". Mol. Microbiol. 54 (2): 298–306. doi:10.1111/j.1365-2958.2004.04275.x. PMID 15469504.
- Bledsoe GH (December 2005). "Malaria primer for clinicians in the United States" (PDF). South. Med. J. 98 (12): 1197–204; quiz 1205, 1230. doi:10.1097/01.smj.0000189904.50838.eb. PMID 16440920.
- Shortt HE (1951). "Life-cycle of the mammalian malaria parasite". Br. Med. Bull. 8 (1): 7–9. PMID 14944807.
History
- Slater LB (2005). "Malarial birds: modeling infectious human disease in animals". Bull Hist Med. 79 (2): 261–94. doi:10.1353/bhm.2005.0092. PMID 15965289.
External links
| Wikispecies has information related to: Plasmodium |
| Wikimedia Commons has media related to Plasmodium. |
- Malaria Atlas Project
- "Plasmodium". NCBI Taxonomy Browser. 5820.