Physcomitrella patens
| Physcomitrella patens | |
|---|---|
 | |
| Scientific classification | |
| Kingdom: | Plantae |
| Division: | Bryophyta |
| Class: | Bryopsida |
| Order: | Funariales |
| Family: | Funariaceae |
| Genus: | Physcomitrella |
| Species: | P. patens |
| Binomial name | |
| Physcomitrella patens (Hedw.) Bruch & Schimp. | |
| Synonyms [1] | |
Physcomitrella patens, the spreading earthmoss,[2] is a moss (bryophyte) used as a model organism for studies on plant evolution, development, and physiology.
Distribution and ecology
Physcomitrella patens is an early colonist of exposed mud and earth around the edges of pools of water.[3][4] P. patens has a disjunct distribution in temperate parts of the world, with the exception of South America.[5] The standard laboratory strain is the "Gransden" isolate, collected by H. Whitehouse from Gransden Wood, in Cambridgeshire.[3]
Model organism
Mosses share fundamental genetic and physiological processes with vascular plants, although the two lineages diverged early in land-plant evolution.[6] A comparative study between modern representatives of the two lines can give an insight into the evolution of the mechanisms behind the complexity of modern plants.[6] In this context, P. patens is used as a model organism.
P. patens is one of a few known multicellular organisms with highly efficient homologous recombination.[7][8] meaning that an exogenous DNA sequence can be targeted to a specific genomic position (a technique called gene targeting) to create knockout mosses. This approach is called reverse genetics and it is a powerful and sensitive tool to study the function of genes and, when combined with studies in higher plants such as Arabidopsis thaliana, can be used to study molecular plant evolution.
The targeted deletion or alteration of moss genes relies on the integration of a short DNA strand at a defined position in the genome of the host cell. Both ends of this DNA strand are engineered to be identical to this specific gene locus. The DNA construct is then incubated with moss protoplasts in the presence of polyethylene glycol. As mosses are haploid organisms, the regenerating moss filaments (protonemata) can be directly assayed for gene targeting within 6 weeks using PCR methods.[9] The first study using knockout moss appeared in 1998 and functionally identified ftsZ as a pivotal gene for the division of an organelle in a eukaryote.[10]
In addition, P. patens is increasingly used in biotechnology. Examples are the identification of moss genes with implications for crop improvement or human health[11] and the safe production of complex biopharmaceuticals in moss bioreactors.[12] By multiple gene knockout Physcomitrella plants were engineered that lack plant-specific post-translational protein glycosylation. These knockout mosses are used to produce complex biopharmaceuticals in a process called molecular farming.[13]
The genome of P. patens, with about 500 megabase pairs organized into 27 chromosomes, was completely sequenced in 2006.[6][14]
Physcomitrella ecotypes, mutants, and transgenics are stored and made freely available to the scientific community by the International Moss Stock Center (IMSC). The accession numbers given by the IMSC can be used for publications to ensure safe deposit of newly described moss materials.
Lifecycle
Like all mosses, the lifecycle of P. patens is characterized by an alternation of two generations: a haploid gametophyte that produces gametes and a diploid sporophyte where haploid spores are produced.
A spore develops into a filamentous structure called protonema, composed of two types of cells – chloronema with large and numerous chloroplasts and caulonema with very fast growth. Protonema filaments grow exclusively by tip growth of their apical cells and can originate side branches from subapical cells. Some side-branch initial cells can differentiate into buds rather than side branches. These buds give rise to gametophores (0.5–5.0 mm[15]), more complex structures bearing leaf-like structures, rhizoids, and the sexual organs: female archegonia and male antheridia. P. patens is monoicous, meaning that male and female organs are produced in the same plant. If water is available, flagellate sperm cells can swim from the antheridia to an archegonium and fertilize the egg within. The resulting diploid zygote originates a sporophyte composed of a foot, seta, and capsule, where thousands of haploid spores are produced by meiosis.
DNA repair and homologous recombination
P. patens is an excellent model in which to analyze repair of DNA damages in plants by the homologous recombination pathway. Failure to repair double-strand breaks and other DNA damages in somatic cells by homologous recombination can lead cell dysfunction or death, and when failure occurs during meiosis, it can cause loss of gametes. The genome sequence of P. patens has revealed the presence of numerous genes that encode proteins necessary for repair of DNA damages by homologous recombination and by other pathways.[6] RpRAD51, a protein at the core of the homologous recombination repair reaction, is required to preserve genome integrity in P. patens.[16] Loss of RpRAD51 causes marked hypersensitivity to the double-strand break-inducing agent bleomycin, indicating that homologous recombination is used for repair of somatic cell DNA damages.[16] RpRAD51 is also essential for resistance to ionizing radiation.[17]
The DNA mismatch repair protein PpMSH2 is a central component of the P. patens mismatch repair pathway that targets base pair mismatches arising during homologous recombination. The PpMsh2 gene is necessary in P. patens to preserve genome integrity.[18] Genes Ppmre11 and Pprad50 of P. patens encode components of the MRN complex, the principal sensor of DNA double-strand breaks.[19] These genes are necessary for accurate homologous recombinational repair of DNA damages in P. patens. Mutant plants defective in either Ppmre11 or Pprad50 exhibit severely restricted growth and development (possibly reflecting accelerated senescence), and enhanced sensitivity to UV-B and bleomycin-induced DNA damage compared to wild-type plants.[19]
 Cleistocarpous sporophyte of the moss P. patens
Cleistocarpous sporophyte of the moss P. patens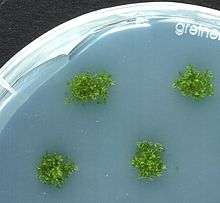
 Moss bioreactor with P. patens
Moss bioreactor with P. patens Four different ecotypes of P. patens stored at the International Moss Stock Center
Four different ecotypes of P. patens stored at the International Moss Stock Center Protonema cells of P. patens
Protonema cells of P. patens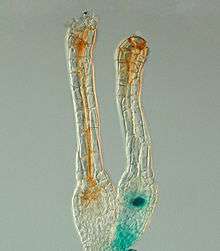 The polycomb gene FIE is expressed (blue) in unfertilised egg cells of the moss P. patens (right) and expression ceases after fertilisation in the developing diploid sporophyte (left). In situ GUS staining of two female sex organs (archegonia) of a transgenic plant expressing a translational fusion of FIE-uidA under control of the native FIE promoter.[20]
The polycomb gene FIE is expressed (blue) in unfertilised egg cells of the moss P. patens (right) and expression ceases after fertilisation in the developing diploid sporophyte (left). In situ GUS staining of two female sex organs (archegonia) of a transgenic plant expressing a translational fusion of FIE-uidA under control of the native FIE promoter.[20] Physcomitrella knockout mutants: Deviating phenotypes induced by transformation with a gene-disruption-library. Physcomitrella wild-type and transformed plants were grown on minimal Knop medium to induce differentiation and development of gametophores. For each plant, an overview (upper row; scale bar corresponds to 1 mm) and a close-up (bottom row; scale bar equals 0.5 mm) are shown. A: Haploid wild-type moss plant completely covered with leafy gametophores and close-up of wild-type leaf. B–D: Different mutants.[21]
Physcomitrella knockout mutants: Deviating phenotypes induced by transformation with a gene-disruption-library. Physcomitrella wild-type and transformed plants were grown on minimal Knop medium to induce differentiation and development of gametophores. For each plant, an overview (upper row; scale bar corresponds to 1 mm) and a close-up (bottom row; scale bar equals 0.5 mm) are shown. A: Haploid wild-type moss plant completely covered with leafy gametophores and close-up of wild-type leaf. B–D: Different mutants.[21]
Taxonomy
| External identifiers for Physcomitrella patens | |
|---|---|
| Encyclopedia of Life | 886166 |
| ITIS | 15827 |
| NCBI | 3218 |
| Also found in: Wikispecies | |
P. patens was first described by Johann Hedwig in his 1801 work Species Muscorum Frondosorum, under the name Phascum patens.[1] Physcomitrella is sometimes treated as a synonym of the genus Aphanorrhegma, in which case P. patens is known as Aphanorrhegma patens.[22] The generic name Physcomitrella implies a resemblance to Physcomitrium, which is named for its large calyptra, unlike that of Physcomitrella.[15]
References
- 1 2 "!Physcomitrella patens (Hedw.) Bruch & Schimp.". Tropicos. Missouri Botanical Garden. Retrieved October 28, 2012.
- ↑ Edwards, Sean R. (2012). English Names for British Bryophytes. British Bryological Society Special Volume. 5 (4 ed.). Wootton, Northampton: British Bryological Society. ISBN 978-0-9561310-2-7. ISSN 0268-8034.
- 1 2 Andrew Cuming (2011). "Molecular bryology: mosses in the genomic era" (PDF). Field Bryology. 103: 9–13.
- ↑ Nick Hodgetts (2010). "Aphanorrhegma patens (Physcomitrella patens), spreading earth-moss". In Ian Atherton; Sam Bosanquet; Mark Lawley. Mosses and Liverworts of Britain and Ireland: a Field Guide (PDF). British Bryological Society. p. 567. ISBN 978-0-9561310-1-0.
- ↑ Stefan A. Rensing, Daniel Lang & Andreas D. Zimmer (2009). Comparative genomics. pp. 42–75. doi:10.1111/b.9781405181891.2009.00003.x. In: Knight et al. (2009).
- 1 2 3 4 Stefan A. Rensing, Daniel Lang, Andreas D. Zimmer, Astrid Terry, Asaf Salamov, Harris Shapiro, Tomoaki Nishiyama, Pierre-François Perroud, Erika A. Lindquist, Yasuko Kamisugi, Takako Tanahashi, Keiko Sakakibara, Tomomichi Fujita, Kazuko Oishi, Tadasu Shin-I, Yoko Kuroki, Atsushi Toyoda, Yutaka Suzuki, Shin-ichi Hashimoto, Kazuo Yamaguchi, Sumio Sugano, Yuji Kohara, Asao Fujiyama, Aldwin Anterola, Setsuyuki Aoki, Neil Ashton, W. Brad Barbazuk, Elizabeth Barker, Jeffrey L. Bennetzen, Robert Blankenship, Sung Hyun Cho, Susan K. Dutcher, Mark Estelle, Jeffrey A. Fawcett, Heidrun Gundlach, Kousuke Hanada, Alexander Heyl, Karen A. Hicks, Jon Hughes, Martin Lohr, Klaus Mayer, Alexander Melkozernov, Takashi Murata, David R. Nelson, Birgit Pils, Michael Prigge, Bernd Reiss, Tanya Renner, Stephane Rombauts, Paul J. Rushton, Anton Sanderfoot, Gabriele Schween, Shin-Han Shiu, Kurt Stueber, Frederica L. Theodoulou, Hank Tu, Yves Van de Peer, Paul J. Verrier, Elizabeth Waters, Andrew Wood, Lixing Yang, David Cove, Andrew C. Cuming, Mitsuyasu Hasebe, Susan Lucas, Brent D. Mishler, Ralf Reski, Igor V. Grigoriev, Ralph S. Quatrano & Jeffrey L. Boore (2008). "The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants". Science. 319 (5859): 64–69. Bibcode:2008Sci...319...64R. doi:10.1126/science.1150646. PMID 18079367.
- ↑ Didier G. Schaefer & Jean-Pierre Zrÿd (1997). "Efficient gene targeting in the moss Physcomitrella patens". Plant Journal. 11 (6): 1195–1206. doi:10.1046/j.1365-313X.1997.11061195.x. PMID 9225463.
- ↑ Didier G. Schaefer (2002). "A new moss genetics: targeted mutagenesis in Physcomitrella patens". Annual Review of Plant Biology. 53: 477–501. doi:10.1146/annurev.arplant.53.100301.135202. PMID 12221986.
- ↑ Annette Hohe, Tanja Egener, JanM. Lucht, Hauke Holtorf, Christina Reinhard, Gabriele Schween & Ralf Reski (2004). "An improved and highly standardised transformation procedure allows efficient production of single and multiple targeted gene-knockouts in a moss, Physcomitrella patens". Current Genetics. 44 (6): 339–347. doi:10.1007/s00294-003-0458-4. PMID 14586556.
- ↑ René Strepp, Sirkka Scholz, Sven Kruse, Volker Speth & Ralf Reski (1998). "Plant nuclear gene knockout reveals a role in plastid division for the homolog of the bacterial cell division protein ftsZ, an ancestral tubulin". Proceedings of the National Academy of Sciences. 95 (8): 4368–4373. Bibcode:1998PNAS...95.4368S. doi:10.1073/pnas.95.8.4368. JSTOR 44902. PMC 22495
 . PMID 9539743.
. PMID 9539743. - ↑ Ralf Reski & Wolfgang Frank (2005). "Moss (Physcomitrella patens) functional genomics – gene discovery and tool development with implications for crop plants and human health". Briefings in Functional Genomics and Proteomics. 4 (1): 48–57. doi:10.1093/bfgp/4.1.48. PMID 15975264.
- ↑ Eva L. Decker & Ralf Reski (2007). "Moss bioreactors producing improved biopharmaceuticals". Current Opinion in Biotechnology. 18 (5): 393–398. doi:10.1016/j.copbio.2007.07.012. PMID 17869503.
- ↑ Anna Koprivova, Christian Stemmer, Friedrich Altmann, Axel Hoffmann, Stanislav Kopriva, Gilbert Gorr, Ralf Reski & Eva L. Decker (2004). "Targeted knockouts of Physcomitrella lacking plant-specific immunogenic N-glycans". Plant Biotechnology Journal. 2 (6): 517–523. doi:10.1111/j.1467-7652.2004.00100.x. PMID 17147624.
- ↑ Ralf Reski, Merle Faust, Xiao-Hui Wang, Michael Wehe & Wolfgang O. Abel (1994). "Genome analysis of the moss Physcomitrella patens (Hedw.) B.S.G.". Molecular and General Genetics. 244 (4): 352–359. doi:10.1007/BF00286686. PMID 8078460.
- 1 2 Bernard Goffinet (2005). "Physcomitrella". Bryophyte Flora of North America, Provisional Publication. Missouri Botanical Garden. Retrieved October 28, 2012.
- 1 2 Markmann-Mulisch U, Wendeler E, Zobell O, Schween G, Steinbiss HH, Reiss B (October 2007). "Differential requirements for RAD51 in Physcomitrella patens and Arabidopsis thaliana development and DNA damage repair". Plant Cell. 19 (10): 3080–9. doi:10.1105/tpc.107.054049. PMC 2174717. PMID 17921313.
- ↑ Schaefer DG, Delacote F, Charlot F, Vrielynck N, Guyon-Debast A, Le Guin S, Neuhaus JM, Doutriaux MP, Nogué F (May 2010). "RAD51 loss of function abolishes gene targeting and de-represses illegitimate integration in the moss Physcomitrella patens". DNA Repair (Amst.). 9 (5): 526–33. doi:10.1016/j.dnarep.2010.02.001. PMID 20189889.
- ↑ Trouiller B, Schaefer DG, Charlot F, Nogué F (2006). "MSH2 is essential for the preservation of genome integrity and prevents homeologous recombination in the moss Physcomitrella patens". Nucleic Acids Res. 34 (1): 232–42. doi:10.1093/nar/gkj423. PMC 1325206. PMID 16397301.
- 1 2 Kamisugi Y, Schaefer DG, Kozak J, Charlot F, Vrielynck N, Holá M, Angelis KJ, Cuming AC, Nogué F (April 2012). "MRE11 and RAD50, but not NBS1, are essential for gene targeting in the moss Physcomitrella patens". Nucleic Acids Res. 40 (8): 3496–510. doi:10.1093/nar/gkr1272. PMC 3333855. PMID 22210882.
- ↑ Assaf Mosquna, Aviva Katz, Eva Decker, Stefan Rensing, Ralf Reski & Nir Ohad (2009). "Regulation of stem cell maintenance by the Polycomb protein FIE has been conserved during land plant evolution". Development. 136: 2433–2444. doi:10.1242/dev.035048. PMID 19542356.
- ↑ Tanja Egener, José Granado, Marie-Christine Guitton, Annette Hohe, Hauke Holtorf, Jan M. Lucht, Stefan A. Rensing, Katja Schlink, Julia Schulte, Gabriele Schween, Susanne Zimmermann, Elke Duwenig, Bodo Rak & Ralf Reski (2002). "High frequency of phenotypic deviations in Physcomitrella patens plants transformed with a gene-disruption library". BMC Plant Biology. 2: 6. doi:10.1186/1471-2229-2-6. PMC 117800. PMID 12123528.
- ↑ Celia Knight, Pierre-François Perroud & David Cove (2009). Preface. pp. xiii–xiv. doi:10.1002/9781444316070.fmatter. In: Knight et al. (2009).
Further reading
- Celia Knight, Pierre-François Perroud & David Cove (2009). The Moss Physcomitrella patens. Annual Plant Reviews. 36. John Wiley & Sons. doi:10.1002/9781444316070. ISBN 978-1-4051-8189-1.
External links
| Wikimedia Commons has media related to Physcomitrella patens. |
- The moss genome consortium homepage
- cosmoss.org - moss transcriptome and genome resource including genome browser
- The Japanese Physcomitrella transcriptome resource (Physcobase)
- The NCBI Physcomitrella patens genome project page
- JGI genome browser
- The moss Physcomitrella patens gives insights into RNA interference in plants
- A small moss turns professional
- Physcomitrella patens facts, developmental stages, organs at GeoChemBio