Photosynthetic reaction centre protein family
| Photosynthetic reaction centre protein | |||||||||
|---|---|---|---|---|---|---|---|---|---|
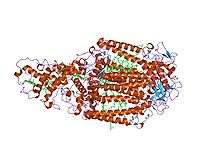 Structure of the photosynthetic reaction centre from Rhodopseudomonas viridis.[1] | |||||||||
| Identifiers | |||||||||
| Symbol | Photo_RC | ||||||||
| Pfam | PF00124 | ||||||||
| InterPro | IPR000484 | ||||||||
| PROSITE | PDOC00217 | ||||||||
| SCOP | 1prc | ||||||||
| SUPERFAMILY | 1prc | ||||||||
| TCDB | 3.E.2 | ||||||||
| OPM superfamily | 2 | ||||||||
| OPM protein | 1dxr | ||||||||
| |||||||||
| Photosynthetic reaction centre cytochrome C subunit | |||||||||
|---|---|---|---|---|---|---|---|---|---|
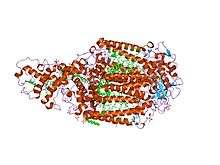 1.96 a x-ray structure of photosynthetic reaction centre from rhodopseudomonas viridis:crystals grown by microfluidic technique | |||||||||
| Identifiers | |||||||||
| Symbol | CytoC_RC | ||||||||
| Pfam | PF02276 | ||||||||
| Pfam clan | CL0317 | ||||||||
| InterPro | IPR003158 | ||||||||
| SCOP | 1prc | ||||||||
| SUPERFAMILY | 1prc | ||||||||
| |||||||||
Photosynthetic reaction centre proteins are main protein components of photosynthetic reaction centres of bacteria and plants.
In bacteria
The photosynthetic apparatus in non-oxygenic bacteria consists of light-harvesting protein-pigment complexes LH1 and LH2, which use carotenoid and bacteriochlorophyll as primary donors.[2] LH1 acts as the energy collection hub, temporarily storing it before its transfer to the photosynthetic reaction centre (RC).[3] Electrons are transferred from the primary donor via an intermediate acceptor (bacteriophaeophytin) to the primary acceptor (quinine Qa), and finally to the secondary acceptor (quinone Qb), resulting in the formation of ubiquinol QbH2. RC uses the excitation energy to shuffle electrons across the membrane, transferring them via ubiquinol to the cytochrome bc1 complex in order to establish a proton gradient across the membrane, which is used by ATP synthetase to form ATP.[4][5][6]
The core complex is anchored in the cell membrane, consisting of one unit of RC surrounded by LH1; in some species there may be additional subunits.[7] RC consists of three subunits: L (light), M (medium), and H (heavy). Subunits L and M provide the scaffolding for the chromophore, while subunit H contains a cytoplasmic domain.[8] In Rhodopseudomonas viridis, there is also a non-membranous tetrahaem cytochrome (4Hcyt) subunit on the periplasmic surface.
In viruses
Photosynthetic reaction centre genes from PSII (PsbA, PsbD) have been discovered within marine bacteriophage.[9][10][11] Though it is widely accepted dogma that arbitrary pieces of DNA can be borne by phage between hosts (transduction), one would hardly expect to find transduced DNA within a large number of viruses. Transduction is presumed to be common in general, but for any single piece of DNA to be routinely transduced would be highly unexpected. Instead, conceptually, a gene routinely found in surveys of viral DNA would have to be a functional element of the virus itself (this does not imply that the gene would not be transferred among hosts - which the photosystem within viruses is[12] - but instead that there is a viral function for the gene, that it is not merely hitchhiking with the virus). However, free viruses lack the machinery needed to support metabolism, let alone photosynthesis. As a result, photosystem genes are not likely to be a functional component of the virus like a capsid protein or tail fibre. Instead, it is expressed within an infected host cell.[13][14] Most virus genes that are expressed in the host context are useful for hijacking the host machinery to produce viruses or for replication of the viral genome. These can include reverse transcriptases, integrases, nucleases or other enzymes. Photosystem components do not fit this mould either. The production of an active photosystem during viral infection provides active photosynthesis to dying cells. This is not viral altruism towards the host, however. The problem with viral infections tends to be that they disable the host relatively rapidly. As protein expression is shunted from the host genome to the viral genome, the photosystem degrades relatively rapidly (due in part to the interaction with light, which is highly corrosive), cutting off the supply of nutrients to the replicating virus.[15] A solution to this problem is to add rapidly degraded photosystem genes to the virus, such that the nutrient flow is uninhibited and more viruses are produced. One would expect that this discovery will lead to other discoveries of a similar nature; that elements of the host metabolism key to viral production and easily damaged during infection are actively replaced or supported by the virus during infection.
Indeed, recently, PSI gene cassettes containing whole gene suites [(psaJF, C, A, B, K, E and D) and (psaD, C, A and B)] were also reported to exist in marine cyanophages from the Pacific and Indian Oceans [16][17][18]
In plant photosystems
This entry describes the photosynthetic reaction centre L and M subunits, and the homologous D1 (PsbA) and D2 (PsbD) photosystem II (PSII) reaction centre proteins from cyanobacteria, algae and plants. The D1 and D2 proteins only show approximately 15% sequence homology with the L and M subunits, however the conserved amino acids correspond to the binding sites of the photochemically active cofactors. As a result, the reaction centres (RCs) of purple photosynthetic bacteria and PSII display considerable structural similarity in terms of cofactor organisation.
The D1 and D2 proteins occur as a heterodimer that form the reaction core of PSII, a multisubunit protein-pigment complex containing over forty different cofactors, which are anchored in the cell membrane in cyanobacteria, and in the thylakoid membrane in algae and plants. Upon absorption of light energy, the D1/D2 heterodimer undergoes charge separation, and the electrons are transferred from the primary donor (chlorophyll a) via phaeophytin to the primary acceptor quinone Qa, then to the secondary acceptor Qb, which like the bacterial system, culminates in the production of ATP. However, PSII has an additional function over the bacterial system. At the oxidising side of PSII, a redox-active residue in the D1 protein reduces P680, the oxidised tyrosine then withdrawing electrons from a manganese cluster, which in turn withdraw electrons from water, leading to the splitting of water and the formation of molecular oxygen. PSII thus provides a source of electrons that can be used by photosystem I to produce the reducing power (NADPH) required to convert CO2 to glucose.[19][20]
Subfamilies
- Photosynthetic reaction centre, M subunit InterPro: IPR005781
- Photosystem II reaction centre protein PsbA/D1 InterPro: IPR005867
- Photosystem II reaction centre protein PsbD/D2 InterPro: IPR005868
- Photosynthetic reaction centre, L subunit InterPro: IPR005871
Notes
- ↑ Deisenhofer J, Epp O, Sinning I, Michel H (February 1995). "Crystallographic refinement at 2.3 A resolution and refined model of the photosynthetic reaction centre from Rhodopseudomonas viridis". J. Mol. Biol. 246 (3): 429–57. doi:10.1006/jmbi.1994.0097. PMID 7877166.
- ↑ Michel H, Oesterhelt D, Lancaster CR, Bibikova MV, Sabatino P (2000). "Structural basis of the drastically increased initial electron transfer rate in the reaction center from a Rh odopseudomonas viridis mutant described at 2.00-A resolution". J. Biol. Chem. 275 (50): 39364–39368. doi:10.1074/jbc.M008225200. PMID 11005826.
- ↑ Hunter CN, Bullough PA, Otto C, Bahatyrova S, Frese RN, Siebert CA, Olsen JD, Van Der Werf KO, Van Grondelle R, Niederman RA (2004). "The native architecture of a photosynthetic membrane". Nature. 430 (7003): –. doi:10.1038/nature02823. PMID 15329728.
- ↑ Scheuring S (2006). "AFM studies of the supramolecular assembly of bacterial photosynthetic core-complexes". Curr Opin Chem Biol. 10 (5): –. doi:10.1016/j.cbpa.2006.08.007. PMID 16931113.
- ↑ Remy A, GerwertK; Gerwert, K (2003). "Coupling of light-induced electron transfer to proton uptake in photosynthesis". Nat. Struct. Biol. 10 (8): 637–644. doi:10.1038/nsb954. PMID 12872158.
- ↑ Michel H, Deisenhofer J (1989). "Nobel lecture. The photosynthetic reaction centre from the purple bacterium Rhodopseudomonas viridis". EMBO J. 8 (8): 2149–2170. PMC 401143
 . PMID 2676514.
. PMID 2676514. - ↑ Miki K, Kobayashi M, Nogi T, Fathir I, Nozawa T (2000). "Crystal structures of photosynthetic reaction center and high-potential iron-sulfur protein from Thermochromatium tepidum: thermostability and electron transfer". Proc. Natl. Acad. Sci. U.S.A. 97 (25): 13561–13566. doi:10.1073/pnas.240224997. PMC 17615. PMID 11095707.
- ↑ Michel H, Ermler U, Schiffer M (1994). "Structure and function of the photosynthetic reaction center from Rhodobacter sphaeroides". J. Bioenerg. Biomembr. 26 (1): 5–15. doi:10.1007/BF00763216. PMID 8027023.
- ↑ Sharon I, Tzahor S, Williamson S, Shmoish M, Man-Aharonovich D, Rusch DB, Yooseph S, Zeidner G, Golden SS, Mackey SR, Adir N, Weingart U, Horn D, Venter JC, Mandel-Gutfreund Y, Béjà O (2007). "Viral photosynthetic reaction center genes and transcripts in the marine environment". ISME J. 1 (6): 492–501. doi:10.1038/ismej.2007.67. PMID 18043651.
- ↑ Millard A, Clokie MR, Shub DA, Mann NH (2004). "Genetic organization of the psbAD region in phages infecting marine Synechococcus strains". Proc. Natl. Acad. Sci. U.S.A. 101 (30): 11007–12. doi:10.1073/pnas.0401478101. PMC 503734. PMID 15263091.
- ↑ Sullivan MB, Lindell D, Lee JA, Thompson LR, Bielawski JP, Chisholm SW (2006). "Prevalence and evolution of core photosystem II genes in marine cyanobacterial viruses and their hosts". PLoS Biol. 4 (8): e234. doi:10.1371/journal.pbio.0040234. PMC 1484495. PMID 16802857.

- ↑ Lindell D, Sullivan MB, Johnson ZI, Tolonen AC, Rohwer F, Chisholm SW (2004). "Transfer of photosynthesis genes to and from Prochlorococcus viruses". Proc. Natl. Acad. Sci. U.S.A. 101 (30): 11013–8. doi:10.1073/pnas.0401526101. PMC 503735. PMID 15256601.
- ↑ Lindell D, Jaffe JD, Johnson ZI, Church GM, Chisholm SW (2005). "Photosynthesis genes in marine viruses yield proteins during host infection". Nature. 438 (7064): 86–9. doi:10.1038/nature04111. PMID 16222247.
- ↑ Clokie MR, Shan J, Bailey S, Jia Y, Krisch HM, West S, Mann NH (2006). "Transcription of a 'photosynthetic' T4-type phage during infection of a marine cyanobacterium". Environ. Microbiol. 8 (5): 827–35. doi:10.1111/j.1462-2920.2005.00969.x. PMID 16623740.
- ↑ Bailey S, Clokie MR, Millard A, Mann NH (2004). "Cyanophage infection and photoinhibition in marine cyanobacteria". Res. Microbiol. 155 (9): 720–5. doi:10.1016/j.resmic.2004.06.002. PMID 15501648.
- ↑ Sharon I, Alperovitch A, Rohwer F, Haynes M, Glaser F, Atamna-Ismaeel N, Pinter RY, Partensky F, Koonin EV, Wolf YI, Nelson N, Béjà O (2009). "Photosystem-I gene cassettes are present in marine virus genomes". Nature. 461 (7261): 258–262. doi:10.1038/nature08284. PMID 19710652.
- ↑ Alperovitch-Lavy A, Sharon I, Rohwer F, Aro EM, Glaser F, Milo R, Nelson N, Béjà O (2011). "Reconstructing a puzzle: existence of cyanophages containing both photosystem-I and photosystem-II gene suites inferred from oceanic metagenomic datasets". Environ. Microbiol. 13 (1): 24–32. doi:10.1111/j.1462-2920.2010.02304.x. PMID 20649642.
- ↑ Béjà O, Fridman S, Glaser F (2012). "Viral clones from the GOS expedition with an unusual photosystem-I gene cassette organization". ISME J. 6 (8): 1617–20. doi:10.1038/ismej.2012.23. PMID 22456446.
- ↑ Kamiya N, Shen JR (2003). "Crystal structure of oxygen-evolving photosystem II from Thermosynechococcus vulcanus at 3.7-A resolution". Proc. Natl. Acad. Sci. U.S.A. 100 (1): 98–103. doi:10.1073/pnas.0135651100. PMC 140893. PMID 12518057.
- ↑ Schroder WP, Shi LX (2004). "The low molecular mass subunits of the photosynthetic supracomplex, photosystem II". Biochim. Biophys. Acta. 1608 (2–3): 75–96. doi:10.1016/j.bbabio.2003.12.004. PMID 14871485.
References
- Deisenhofer, J.; Epp, O.; Miki, K.; Huber, R.; Michel, H. (1984). "X-ray structure analysis of a membrane protein complex. Electron density map at 3 a resolution and a model of the chromophores of the photosynthetic reaction center from Rhodopseudomonas viridis". Journal of Molecular Biology. 180 (2): 385–398. PMID 6392571.