Neurotransmission
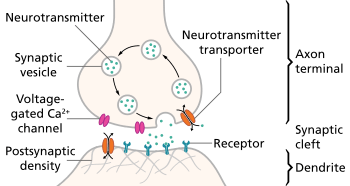
Neurotransmission (Latin: transmissio "passage, crossing" from transmittere "send, let through"), also called synaptic transmission, is the process by which signaling molecules called neurotransmitters are released by a neuron (the presynaptic neuron), and bind to and activate the receptors of another neuron (the postsynaptic neuron). Neurotransmission is essential for the process of communication between two neurons. Synaptic transmission relies on: the availability of the neurotransmitter; the release of the neurotransmitter by exocytosis; the binding of the postsynaptic receptor by the neurotransmitter; the functional response of the postsynaptic cell; and the subsequent removal or deactivation of the neurotransmitter.
In response to a threshold action potential or graded electrical potential, a neurotransmitter is released at the presynaptic terminal. The released neurotransmitter may then move across the synapse to be detected by and bind with receptors in the postsynaptic neuron. Binding of neurotransmitters may influence the postsynaptic neuron in either an inhibitory or excitatory way. The binding of neurotransmitters to receptors in the postsynaptic neuron can trigger either short term changes, such as changes in the membrane potential called postsynaptic potentials, or longer term changes by the activation of signaling cascades.
Neurons form elaborate networks through which nerve impulses (action potentials) travel. Each neuron has as many as 15,000 connections with other neurons. Neurons do not touch each other (except in the case of an electrical synapse through a gap junction); instead, neurons interact at close contact points called synapses. A neuron transports its information by way of an action potential. When the nerve impulse arrives at the synapse, it may cause the release of neurotransmitters, which influence another (postsynaptic) neuron. The postsynaptic neuron may receive inputs from many additional neurons, both excitatory and inhibitory. The excitatory and inhibitory influences are summed, and if the net effect is inhibitory, the neuron will be less likely to "fire" (i.e., generate an action potential), and if the net effect is excitatory, the neuron will be more likely to fire. How likely a neuron is to fire depends on how far its membrane potential is from the threshold potential, the voltage at which an action potential is triggered because enough voltage-dependent sodium channels are activated so that the net inward sodium current exceeds all outward currents.[1] Excitatory inputs bring a neuron closer to threshold, while inhibitory inputs bring the neuron farther from threshold. An action potential is an "all-or-none" event; neurons whose membranes have not reached threshold will not fire, while those that do must fire. Once the action potential is initiated (traditionally at the axon hillock), it will propagate along the axon, leading to release of neurotransmitters at the synaptic bouton to pass along information to yet another adjacent neuron.
Stages in neurotransmission at the synapse
- Synthesis of the neurotransmitter. This can take place in the cell body, in the axon, or in the axon terminal.
- Storage of the neurotransmitter in storage granules or vesicles in the axon terminal.
- Calcium enters the axon terminal during an action potential, causing release of the neurotransmitter into the synaptic cleft.
- After its release, the transmitter binds to and activates a receptor in the postsynaptic membrane.
- Deactivation of the neurotransmitter. The neurotransmitter is either destroyed enzymatically, or taken back into the terminal from which it came, where it can be reused, or degraded and removed.[2]
Summation
Each neuron connects with numerous other neurons, receiving numerous impulses from them. Summation is the adding together of these impulses at the axon hillock. If the neuron only gets excitatory impulses, it will generate an action potential. If instead the neuron gets as many inhibitory as excitatory impulses, the inhibition cancels out the excitation and the nerve impulse will stop there.[3] Action potential generation is proportionate to the probability and pattern of neurotransmitter release, and to postsynaptic receptor sensitization.[4][5][6]
Spatial summation means that the effects of impulses received at different places on the neuron add up, so that the neuron may fire when such impulses are received simultaneously, even if each impulse on its own would not be sufficient to cause firing.
Temporal summation means that the effects of impulses received at the same place can add up if the impulses are received in close temporal succession. Thus the neuron may fire when multiple impulses are received, even if each impulse on its own would not be sufficient to cause firing.[7]
Convergence and divergence
Neurotransmission implies both a convergence and a divergence of information. First one neuron is influenced by many others, resulting in a convergence of input. When the neuron fires, the signal is sent to many other neurons, resulting in a divergence of output. Many other neurons are influenced by this neuron.[8]
Cotransmission
Cotransmission is the release of several types of neurotransmitters from a single nerve terminal.
At the nerve terminal, neurotransmitters are present within 35–50 nm membrane-encased vesicles called synaptic vesicles. To release neurotransmitters, the synaptic vesicles transiently dock and fuse at the base of specialized 10–15 nm cup-shaped lipoprotein structures at the presynaptic membrane called porosomes.[9] The neuronal porosome proteome has been solved, providing the molecular architecture and the complete composition of the machinery.[10]
Recent studies in a myriad of systems have shown that most, if not all, neurons release several different chemical messengers.[11] Cotransmission allows for more complex effects at postsynaptic receptors, and thus allows for more complex communication to occur between neurons.
In modern neuroscience, neurons are often classified by their cotransmitter. For example, striatal "GABAergic neurons" utilize opioid peptides or substance P as their primary cotransmitter.
Some neurons can release at least two neurotransmitters at the same time, the other being a cotransmitter, in order to provide the stabilizing negative feedback required for meaningful encoding, in the absence of inhibitory interneurons.[12] Examples include:
- GABA–glycine co-release.
- Dopamine–glutamate co-release.
- Acetylcholine (Ach)–glutamate co-release.
- ACh–vasoactive intestinal peptide (VIP) co-release.
- ACh–calcitonin gene-related peptide (CGRP) co-release.
- Glutamate–dynorphin co-release (in hippocampus).
See also
- Autoreceptor
- Biological neuron model § Synaptic transmission
- G protein-coupled receptor
- Molecular neuropharmacology
- Neuromuscular transmission
- Neuropsychopharmacology
- Neurotransmitter
References
- ↑ Holden, Arun; Winlow, William (1984). The Neurobiology of Pain: Symposium of the Northern Neurobiology Group Held at Leeds on 18 April 1983 (1st ed.). Manchester Univ Pr. p. 111. ISBN 0719010616.
- ↑ Kolb, Bryan; Whishaw, Ian Q. (2003). Fundamentals of Human Neuropsychology (5th ed.). Worth. pp. 102–104. ISBN 978-0-7167-5300-1. (reference for all five stages)
- ↑ Purves D, Augustine GJ, Fitzpatrick D, et al., eds. (2001). Summation of Synaptic Potentials. Neuroscience (2nd ed.). Sunderland (MA): Sinauer Associates.
- ↑ Jin-Hui Wang; J. Wei; X. Chen; J. Yu; N. Chen; J. Shi (2008). "Gain and fidelity of transmission patterns at cortical excitatory unitary synapses improve spike encoding". Journal of Cell Science. 121 (17): 2951–2960. doi:10.1242/jcs.025684. PMID 18697836.
- ↑ Jiandong Yu; Hao Qian; Na Chen; Jin-Hui Wang (2011). "Quantal glutamate release is essential for reliable neuronal encodings in cerebral networks". PloS One. 6 (9): e25219. doi:10.1371/journal.pone.0025219. PMID 21949885.
- ↑ Jiandong Yu; Hao Qian; Jin-Hui Wang (2012). "Upregulation of transmitter release probability improves a conversion of synaptic analogue signals into neuronal digital spikes". Molecular Brain. 5 (26): 1–11. doi:10.1186/1756-6606-5-26. PMC 3497613
 . PMID 22852823.
. PMID 22852823. - ↑ Archived February 19, 2006, at the Wayback Machine.
- ↑ Archived September 13, 2006, at the Wayback Machine.
- ↑ Anderson, L. L. (2006). "Discovery of the 'porosome' The universal secretory machinery in cells". Journal of Cellular and Molecular Medicine. 10 (1): 126–31. doi:10.1111/j.1582-4934.2006.tb00294.x. PMID 16563225.
- ↑ Lee, Jin-Sook; Jeremic, Aleksandar; Shin, Leah; Cho, Won Jin; Chen, Xuequn; Jena, Bhanu P. (2012). "Neuronal porosome proteome: Molecular dynamics and architecture". Journal of Proteomics. 75 (13): 3952–62. doi:10.1016/j.jprot.2012.05.017. PMID 22659300.
- ↑ Trudeau, L.-E.; Gutiérrez, R. (2007). "On Cotransmission & Neurotransmitter Phenotype Plasticity". Molecular Interventions. 7 (3): 138–46. doi:10.1124/mi.7.3.5. PMID 17609520.
- ↑ Thomas, E.A.; Bornstein, J.C. (2003). "Inhibitory cotransmission or after-hyperpolarizing potentials can regulate firing in recurrent networks with excitatory metabotropic transmission". Neuroscience. 120 (2): 333–51. doi:10.1016/S0306-4522(03)00039-3. PMID 12890506.
{kind=link}
{kind=link}