Neuroscience and sexual orientation
| Sexual orientation |
|---|
| Sexual orientations |
| Non-binary categories |
| Research |
| Non-human animals |
|
Sexual orientation is an enduring pattern of romantic or sexual attraction (or a combination of these) to persons of the opposite sex or gender, the same sex or gender, or to both sexes or more than one gender.[1][2] The ultimate causes and mechanisms of sexual orientation development in humans remain unclear and many theories are speculative and controversial. However, advances in neuroscience explain and illustrate characteristics linked to sexual orientation. Studies have explored structural neural-correlates, functional and/or cognitive relationships, and developmental theories relating to sexual orientation in humans.
Developmental neurobiology
Many theories concerning the development of sexual orientation involve fetal neural development, with proposed models illustrating prenatal hormone exposure, maternal immunity, and developmental instability. Other proposed factors include genetic control of sexual orientation. No conclusive evidence has been shown that environmental or learned effects are responsible for the development of non-heterosexual orientation.[3]
Prenatal androgen model
Sexual dimorphisms in the brain and behavior among vertebrates are accounted for by the influence of gonadal steroidal androgens as demonstrated in animal models over the past few decades. The prenatal androgen model of homosexuality describes the neuro-developmental effects of fetal exposure to these hormones.[3] In 1985, Geschwind and Galaburda proposed that homosexual men are exposed to high androgen levels early in development, explaining their tendency to be less right-handed and by extension the hyper-masculinized traits observed in this population.[3] It is currently argued that temporal and local variations in androgen exposure to a fetus’s developing brain is a factor in the pathways determining homosexuality. Recently research has moved on to finding somatic markers for prenatal hormonal exposure that can be easily, and non-invasively, explored in otherwise endocrinologically normal populations. As these somatic markers are known to be influenced by prenatal sex hormones, showing variation in these features between homosexuals and heterosexuals can provide a ‘window’ into the early neurodevelopment of sexual orientation under the influence of prenatal hormones. Various somatic markers (including 2D:4D finger ratios, auditory evoked potentials, fingerprint patterns and eye-blink patterns) have since been found to show variation based on sexual orientation in healthy adult individuals.[3][4]
Other evidence supporting the role of testosterone and prenatal hormones in sexual orientation development include observations of male subjects with cloacal exstrophy who were sex-assigned as female during birth only later to declare themselves male. This supports the theory that the prenatal testosterone surge is crucial for gender identity development. Additionally, females whose mothers were exposed to diethylstilbestrol (DES) during pregnancy show higher rates of bi- and homosexuality.[5]
2D:4D digit ratio
The best non-invasive marker of prenatal hormone exposure is the digit ratio of the second and fourth finger lengths (2D:4D ratio), a known sexually dimorphic measure (males showing lower ratios than females). Patients with androgen over-exposure (such as in congenital adrenal hyperplasia) show lower 2D:4D ratios,[6][7] providing evidence linking prenatal androgen exposure as key to this feature. XY individuals with androgen insensitivity syndrome due to a dysfunctional gene for the androgen receptor present as women and have feminine digit ratios, as would be predicted if androgenic hormones affect digit ratios. This finding also demonstrates that the sex difference in digit ratio is unrelated to the Y chromosome per se.[8] Additionally, the 2D:4D ratio has been shown to be affected by variation in the androgen receptor gene in men.[9] The ratio of testosterone to estrogen in amniotic fluid has also been found to be negatively correlated with the 2D:4D ratio.[3]
Independent studies indicate that homosexual women have masculinized (lower) digit ratios,[10][11][12][13][14][15][16][17][18][19] and homosexual men show either hyper-masculinized or feminized ratios. These findings reinforce the prenatal androgen model that non-typical prenatal hormone exposure is related to the development of human homosexuality.[3]
Auditory evoked potentials
Studies of the central nervous system processing of auditory sensation, aspects of which has been linked to prenatal androgen exposure, to click-stimuli have shown that homosexual women have masculinized responses while homosexual men have hyper-masculinized responses.[3]
Fraternal birth order effect
Studies show that homosexual men have higher numbers of older brothers than heterosexual men.[20] This finding led to the discovery of the fraternal birth order effect, according to which the more older brothers a man has from the same mother, the greater the probability is that he will have a homosexual orientation. Estimations indicate that there is a 33% increase in chances of homosexuality in a male child with each older brother.[3] The fraternal birth order effect holds true only for biological brothers and the chances of male homosexuality is not increased by the number of older stepbrothers or adopted siblings.[21] It is estimated that one-seventh of all homosexual males owe their sexual orientation to the fraternal birth order effect.[22][23][24] The effect only applies to right-handed homosexual males; it does not increase the likelihood of homosexuality in left-handed or ambidextrous males.[25] As the effect is contingent on handedness and handedness is a prenatally determined trait, the fraternal birth order effect is understood to be biological, rather than psychosocial, in nature and is known to operate prenatally.[21]
The exact biological mechanism by which the effect operates during prenatal life is currently unconfirmed. The proposed mechanism by which the effect is believed to operate states that a mother develops an immune response against a substance important in male fetal development during pregnancy, and that this immune effect becomes increasingly likely with each male fetus gestated by the mother. This immune effect is thought to cause an alteration in (some) later born males’ prenatal brain development. The target of the immune response may be molecules (i.e., Y-linked proteins) on the surface of male fetal brain cells, including in sites of the anterior hypothalamus (which has been linked to sexual orientation in other research). Antibodies produced during the immune response are thought to cross the placental barrier and enter the fetal compartment where they bind to the Y-linked molecules and thus alter their role in sexual differentiation, leading some later born males to be attracted to men as opposed to women. To date, the proposed mechanism has only indirect evidence to support it. It is also the only plausible mechanism proposed so far to explain how the fraternal birth order effect may operate in utero.[21] The fraternal birth order effect does not apply to the development of female homosexuality.[21]
Developmental instability and handedness
Developmental instability refers to an organism’s degree of vulnerability to environmental and genetic stresses during development.[3][26][27] Measures of this instability provide some insight into the developmental history of the organism. Asymmetry in bilateral features of the body, known as fluctuating asymmetry, is often employed in research as a proxy measure of developmental instability. Consistent non-right handedness is also said to constitute a measure of developmental instability.[26]
The chances of being left-handed may be increased in homosexual populations. In comparison with a heterosexual sample, a 2000 meta-analysis of earlier studies by Lalumiere et al.[27] showed that homosexual men have approximately one-third (34%) higher odds of being left-handed while homosexual women have almost twice (91%) higher odds of being so.[27] It has been proposed that non-right-handedness (including ambidexterity) is related to homosexuality through developmental instability.[25] According to this hypothesis, same-sex orientation is due to generalized developmental factors (i.e., non-hormonal environmental or genetic factors) that shift erotic preferences away from the species-typical pattern of opposite-sex attraction during the neurodevelopment of the fetus. If this logic is correct, one would expect that male and female homosexuality should be associated with other signs of developmental instability such as increased fluctuating asymmetry (e.g., in dermatoglyphics, and the lengths of ears, wrists, fingers and feet).[3][26]
However, several studies have found no significant differences in fluctuating asymmetry between heterosexuals and homosexuals,[3][14][28][29] suggesting that a homosexual orientation is not a ‘less than optimal’ phenotypic sexual orientation.[3] On the contrary, it has been found that gay men and lesbian women actually show less fluctuating asymmetry (and thus, less developmental instability) than heterosexual men or women, not more. In other words, heterosexuals have a weaker genomic ability than homosexuals to successfully buffer development to achieve a normal phenotype under imperfect environmental conditions.[30][31] Additionally, it has been found that heterosexual male-to-female (MtF) transsexuals (i.e., MtF transsexuals who are attracted to women) exhibited greatest fluctuating asymmetry in comparison to homosexual and control groups, which shows that developmental instability may account for variations in gender identity but not sexual orientation per se.[26] It has also been found that homosexual men are stereotypically considered more attractive than heterosexual men, homosexual men are actually rated as more attractive than heterosexual men (even when the raters do not know the men's sexual orientations), and in childhood, independent raters describe gender atypical boys as more attractive than gender-typical boys.[26][32][33][34] Thus, something in the physiognomy of homosexual children and adults marks them out as more attractive. It has been suggested that this is due to the fact that homosexual men have low fluctuating asymmetry, which is known to be associated with greater attractiveness.[26][35][36]
Hence, the general mechanism proposed by the developmental instability account of Lalumiere et al. for both male and female homosexuality is incompatible with the variance suggested by the evidence for different pathways to male versus female homosexuality.[3][37] The developmental instability hypothesis has also been criticized for being too domain general and for being unclear on exactly what developmental mechanisms are supposedly disrupted.[26] Thus, the canalization of the sexual orientation trait is more likely due to specific neurodevelopmental mechanisms (which may include the actions of prenatal androgens) rather than general-purpose neurodevelopmental mechanisms (such as developmental instability).[3]
A significant problem with the measurement of fluctuating asymmetry is that many of the anatomical features that are used for the measurement of fluctuating asymmetry also display some degree of directional asymmetry (i.e., developmentally normal deviation from symmetry that is in the same direction in most individuals, such as the position of the heart on the left side of the chest), and the direction and magnitude of the directional asymmetry can vary between the sexes and between homosexuals and heterosexuals (homosexuals have comparatively more directional asymmetry than heterosexuals).[14][30] If this directional asymmetry is not carefully assessed and removed from the data for each group of subjects, the measure of fluctuating asymmetry is likely to be incorrect. When datasets from previously published studies (that appeared to show increased fluctuating asymmetry in homosexuals) were reanalyzed in light of this issue, it was found that homosexual men and women actually show less fluctuating asymmetry than heterosexual men or women. This suggests that homosexual people experience less developmental instability than heterosexual people.[30]
Structural differences
Postmortem and imaging studies over the past two decades have revealed structural differences in both global structures and sexually-related brain structures between heterosexual and homosexual subjects.
Hypothalamus
The hypothalamus is a portion of the brain that contains a number of 'nuclei' (discrete groups of cell bodies of neurons).[note 1] The hypothalamus is known to be involved in sex differences in reproductive behavior, mediating responses in menstrual cycles in women and specifically the anterior hypothalamus of the brain helps regulate male-typical sexual behavior. Recently, the hypothalamus has been linked to gender identity and sexual orientation.[40]
A seminal paper by Simon LeVay found that an interstitial nucleus of the hypothalamus, INAH3, was dimorphic according to sexual orientation not gender. Specifically, the INAH3 of homosexual men was found to be smaller in volume than that of heterosexual men. These results were obtained from postmortem analysis of hypothalamic nuclei of known homosexual subjects compared to heterosexual patients.[41] Further research has found that the INAH3 is of smaller volume in homosexual men than in heterosexual men because homosexual men have a higher neuronal density in the INAH3 than heterosexual men; there is no difference in the number or cross-sectional area of neurons in the INAH3 of homosexual versus heterosexual men.[42] It has also been found that there is no effect of HIV infection on the size of INAH3, that is, HIV infection does not account for the observed difference in INAH3 volume between homosexual and heterosexual men.[42]
The hypothalamus is also linked to sexual orientation through findings that show that activity of aromatase, an important enzyme converting androgens to estrogens, is high in the preoptic hypothalamic region of mammals during the pre- and neonatal periods. This activity is linked to sexual differentiation and may be a basis in structural and functional sexual differences playing a role in mediating the sexual orientation development due to prenatal hormonal exposure.[40]
The suprachiasmatic nucleus (SCN) of the anterior hypothalamus has also been found to relate to sexual orientation, as it is larger in size and more elongated in shape in homosexual males than in heterosexual males and females. The vasopressin-containing subnucleus of the SCN of homosexual men is twice as large and has 2.1 times as many cells than the vasopressin-containing subnucleus of the SCN in heterosexual men.[40][43] This may be a neurological explanation for the finding that homosexual men arise and retire earlier each day than heterosexuals, as it is known that the SCN is involved in modulating human circadian rhythms.[40] Analogously, in a rat model study, it was found that male rats treated with an aromatase inhibitor showed a partner preference for females when tested in the late dark phase but showed homosexual mating preferences when tested in the early dark phase, implicating the involvement of the SCN in sexual orientation in other species.[40]
 Location of the hypothalamus in the brain. (mid-sagittal view)
Location of the hypothalamus in the brain. (mid-sagittal view) Location of the SCN (green) within the hypothalamus (blue). (mid-sagittal view)
Location of the SCN (green) within the hypothalamus (blue). (mid-sagittal view)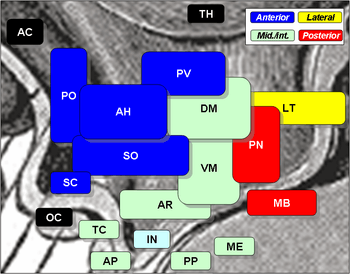 Locations of various hypothalamic nuclei (mid-sagittal view). The SCN is labelled as 'SC' in blue. The INAH3 is not shown.
Locations of various hypothalamic nuclei (mid-sagittal view). The SCN is labelled as 'SC' in blue. The INAH3 is not shown. A cross section of the hypothalamus displays the SCN on either side of the brain's third ventricle. (frontal view)
A cross section of the hypothalamus displays the SCN on either side of the brain's third ventricle. (frontal view)
Thalamus
The thalamus is a midline symmetrical ovoid structure within the human brain, situated between the cerebral cortex and the midbrain in both cerebral hemispheres. An MRI study compared subcortical volumes of homosexual men and heterosexual men. It found that while both groups did not differ in total brain volume, the volume of the thalamus (in both cerebral hemispheres) was larger in heterosexual men than in homosexual men.[44] Another study reported that functional connectivity involving the right thalamus and the right cuneus were different between homosexual and heterosexual men, and even showed correlations with Kinsey scale scores.[45] Additionally, the thalamus is reportedly involved in sexual arousal and reward processes; during visually evoked sexual arousal both heterosexual men and homosexual men activated the thalamus, but in contrast to homosexual men, heterosexual men showed additional activation in the lingual gyrus.[44]
Bed nucleus of the stria terminalis
The bed nucleus of the stria terminalis (BNST) is a forebrain limbic area that is involved in control of mating behaviour. It receives neuronal inputs from the medial amygdala and the accessory olfactory bulb and sends projections to both the medial preoptic area and the ventromedial nucleus of the hypothalamus.[46][47] The central part of the BNST (BNSTc) is 44% larger in heterosexual men than heterosexual women and 62% larger in homosexual men than heterosexual women.[48] The BNSTc is larger in homosexual men than heterosexual men, though the difference in size is not statistically significant.[48] The BNSTc of homosexual men is said to be ‘hypermasculinised’ as it is larger than the BNSTc of both heterosexual men and women.[47]
Amygdala
It has been found that both homosexual men and homosexual women show different amygdala connections than do heterosexual men and women. Specifically, the amygdala connections of homosexual men and heterosexual women were found to be more widespread from the left amygdala, whereas in heterosexual men and homosexual women the functional connections were more widespread from the right amygdala.[49][50]
Anterior commissure
The anterior commissure, a bundle of white matter fibers connecting the two cerebral hemispheres, was found by Allen and Gorski to be larger in homosexual men and heterosexual women than in heterosexual men.[40] This finding provides a possible anatomical basis for higher inter-hemispheric functional connections in homosexuals explaining why homosexual men and heterosexual women show language circuit functional symmetry in out performing heterosexual men in verbal tests.[49]

Corpus callosum
The corpus callosum (CC), like the anterior commissure, is a major neuronal link connecting the two cerebral hemispheres. However, unlike the anterior commissure (which is present in all vertebrates), the CC is only present in placental animals (including humans).[52] An MRI study comparing the CC of right-handed homosexual and heterosexual men found that all parts of the CC are larger in homosexual men.[53] In particular, the isthmus (a part of the CC between the callosal body and the splenium) is significantly larger in size in homosexual than heterosexual men.[51][52] The size of the CC has a strong genetic basis, with heritability rates between 82–94%.[51] This association of sexual orientation with a brain structure having high heritability supports a genetic and neurobiological basis in the origin of sexual orientation.[51]
Gray matter
Gray matter is a major part of the central nervous system that is composed mostly of neuronal cell bodies. While men generally have greater amounts of grey and white matter than women (due to men’s larger body mass and consequently larger brain size), women generally possess a greater grey matter-to-white matter ratio and thicker layers of grey matter in specific cortical areas compared to men.[54][55] It has been found that homosexual women have relatively less gray matter than heterosexual women in the ventral cerebellum, left ventral premotor cortex, temporo-basal cortex, and most significantly, in the left perirhinal cortex. No difference in amount of gray matter was found between heterosexual and homosexual men.[54]
These findings are important because the perirhinal cortex is located near brain regions (entorhinal cortex, hippocampus, parahippocampal gyrus, and amygdala) involved in olfactory and spatial processing, which have been shown to exhibit differences in sexual orientation — specifically, homosexual women are known to perform higher than heterosexual women in spatial processing tests.[50][54] The perirhinal cortex itself is involved in functions related to the processing of sexual stimuli such as olfactory processing, memory encoding and spatial processing. It is also involved in detection of object identity. Olfactory processing is known to modify sexual attraction in humans and the olfactory system is able to differentiate pheromone-like compounds according to sexual orientation.[54]
The finding that homosexual women have a “male-like” GM pattern but homosexual men do not have a “female-like” pattern indicates that male and female homosexuality do not manifest the same way at a structural level in the brain. In addition, other findings of sexually dimorphic features that are more male-like in homosexual women, but not female-like in homosexual men, include otoacoustic emissions, the 2D:4D finger ratio, and body build. Altogether, these findings suggest that sex-atypical levels of prenatal androgen action may be involved in the origin of female homosexuality.[54]
Cerebral asymmetry
The size of the brain’s hemispheres is a sexually dimorphic trait in which men tend to show asymmetry in the volumes of their hemispheres while women show volumetric symmetry. It is also a trait that is unlikely to be affected by any learned patterns. A 2008 volumetric MRI study indicated that homosexual men and heterosexual women showed hemispheric volumes to be symmetric while homosexual women and heterosexual men showed rightward asymmetry in hemispheric volumes. These findings demonstrate a global neurological difference in brain structures showing sex-atypical characteristics associated with sexual orientation.[49][50]
Cerebral cortex
The cerebral cortex is the human cerebrum's outer layer and is composed of neural tissue. An MRI study compared cortical thickness in various brain regions of homosexual men, heterosexual men and heterosexual women. It found that homosexual men had thinner cortices compared to heterosexual men in the right hemisphere's lateral orbitofrontal region, as well as in regions located in the visual cortex (lingual, pericalcarine and cuneus). The same regions also showed thinner cortex in heterosexual women than in heterosexual men, whereas no differences were found between heterosexual women and homosexual men. Homosexual and heterosexual men did not differ in total brain volumes and it was determined that the reported differences in cortical thickness were not affected by the subjects’ years of education or brain volume. As the aforementioned regions exhibit sexual dimorphism, the authors speculated that biological processes frequently proposed to underlay sexual dimorphism, such as gene-dependent and sex hormone-dependent mechanisms during prenatal and postnatal development, may interact with the cortical architecture in visual areas resulting in different cortical thickness in homosexual versus heterosexual men.[44]
Additionally, homosexual men showed thinner cortices than both heterosexual men and women in the right pars triangularis (also known as Brodmann area 45) and inferior temporal regions. This suggests that cerebral differences related to male homosexuality may also be present in regions which are not necessarily considered as sexually dimorphic.[44][note 2]
Functional differences
Recent studies have begun exploring the functional and cognitive substrates of sexual orientation, ultimately a behavioral manifestation. Neural processing in response to specific stimuli and sexually-biased cognitive tasks have been found to be correlated with an individual’s sexual orientation.
Response to pheromones
Two proposed human pheromones – the progesterone derivative 4,16-androstadien-3-one (AND) and an estrogen-like steroid estra-1,3-5(10),16-tetraen-3-ol (EST) – have been shown to have sexual orientation specific responses in activating the neural circuits of the anterior hypothalamus in homosexual and heterosexual subjects. The anterior hypothalamus is involved in processing reproductive functions and recent evidence suggests it helps integrate hormonal and sensory cues involved in sexual behavior and sexual preference.[57]
Recent functional neuro-imaging experiments have shown that the presentation of AND, found in male sweat, as an olfactory stimulus produced normal olfactory responses in heterosexual men and homosexual women, while activating the anterior hypothalamus in homosexual men and heterosexual women.[58] The proposed pheromone EST, found in the urine of pregnant women, produces normal olfactory activation in homosexual men and heterosexual women while homosexual women and heterosexual men demonstrated sexually-related hypothalamic responses.[57]
Homosexual men showed the same sexually-related functional responses to these stimuli as heterosexual women and homosexual women responded like heterosexual men. This research conducted by Berglund and Savic indicates overall that AND and EST induce “sex-specific effects on the autonomic nervous system” and that the stimuli elicited a response pathway that was dependent on the subject’s sexual orientation rather than phenotypic sex.[58]
Response to visual sexual stimuli
Sexual arousal is a highly coordinated process that prepares a person for reproductive behavior. Widespread changes occur in the person’s neurophysiological state during arousal to achieve adaptive responses. The person’s attentive, affective, and motivational systems are optimized so that successful selection and engagement of sexual stimuli may occur. In response to visual sexual stimuli, men exhibit category-specific genital and self-reported subjective sexual arousal. Their greatest sexual arousal is to the categories of people with whom they prefer to have sex — homosexual men experience higher genital and subjective arousal to men than to women (and thus prefer male sexual stimuli) whereas heterosexual men experience higher genital and subjective arousal to women than to men (and thus prefer female sexual stimuli). Hormones are believed to prenatally influence the development of neural structures that regulate sexual behavior. Hence, it is thought that certain aspects of neurohormonal development in homosexuals proceed in a manner different from heterosexuals, resulting in psychological differences such as distinct triggers (or 'stimuli') for sexual arousal.[59]
A 2007 functional magnetic resonance imaging (fMRI) study exploring the neural mechanisms of sexual arousal in homosexual and heterosexual men showed their subjects male–male and female–female sexual interactions. They demonstrated that homosexual and heterosexual men activate the same brain regions after each one is exposed to a sexual stimulus concordant with the subject’s sexual orientation (i.e., male–male sexual interactions for homosexual men and female–female sexual interactions for heterosexual men).[59]
Another fMRI study demonstrated that upon viewing of both male–female and male–male erotic visual stimuli, only those videos corresponding to the subject’s sexual orientation produced activation patterns in brain areas associated with sexual arousal. The response of heterosexuals viewing male–female adult videos showed the same pattern of sexual arousing neural processing as homosexuals viewing male–male adult videos, while the viewing of the opposite orientation’s images did not elicit the same response. Significant correlation was found between sexual arousal and neural activation in the hypothalamus, a key brain region in sexual function. Self-reported sexual arousal ratings were also equal in both groups. However, the magnitude of hypothalamic activation was lower in homosexual men than heterosexual men, a characteristic that is shared by women who are attracted to men.[60]
Another fMRI study determined the patterns of brain activation in homosexual and heterosexual male subjects while exposing them to male–male, male–female and female–female sexual visual stimuli. They found that different neural circuits were active in homosexual and heterosexual men: brain regions such as the left angular gyrus, right pallidum and left caudate nucleus were exclusively activated in homosexual men whereas the bilateral lingual gyrus, the right parahippocampal gyrus and the right hippocampus were exclusively activated in heterosexual men. These findings indicate that the neural circuits (related to the processing of visual sexual stimuli) that are active during sexual arousal in homosexual and heterosexual men are different.[61]
Another fMRI study showed heterosexual and homosexual men and women photographs of aroused male and female genitals. By limiting the visual sexual stimulus to photographs of genitals, the authors minimized neuronal activity related to neuronal processing of various stimuli such as faces, voices, body movements, and sexually arousing body parts other than genitals. They found that the ventral striatum, centromedian thalamus and bilateral ventral premotor cortex showed a stronger response to photographs of aroused genitals of the preferred sex relative to corresponding photographs of the non-preferred sex. As the ventral striatum and centromedian thalamus are known to be activated by innate preferences, the selective responsiveness of these regions to preferred sexual stimuli appears to reflect a predetermined response pattern. This notion is supported by evidence that sexual orientation is biological in origin.[62]
Another fMRI study sought to test whether subjects would respond more to faces of the sex (male or female) they were sexually oriented towards and predicted such modulation in the brain's reward circuitry. Heterosexual and homosexual men and women were shown photographs of male and female faces and were asked to assess facial attractiveness. Consistent with the hypothesis, it was found that the reward circuitry of homosexual males and heterosexual females responded more to photographs of male faces, whereas the reward circuitry of homosexual females and heterosexual males responded more to photographs showing female faces. The interaction between stimulus gender (male or female face) and the sexual orientation (homosexual or heterosexual) of the subject was highly significant in two brain regions: the mediodorsal nucleus of the thalamus (mdT) and the medial orbitofrontal cortex (OFC). Activation in the OFC is notable because the OFC is involved in representing the reward value of various sensory stimuli, including attractive faces. It also appears that the OFC has an important role in the processing of facial cues required for social communication, as this region possesses face-selective neurons and because patients with OFC lesions are unable to identify emotional facial expressions. The modulation of the response to faces within the OFC by sexual orientation further adds to its role in social behavior. Since the mdT and OFC receive neural projections from each other, the similar patterns of activation observed in these regions can be attributed to their anatomical connections.[63]
Functional cerebral asymmetry
Differences in neural processing and cognitive tasks have been found in relation to sexual orientation. In a 1987 review on cognition, cerebral lateralization, and sexual orientation, Sanders and Ross-Field suggested that prenatal hormonal events would lead to functional cerebral asymmetries related to sexual orientation.[64]
Certain cognitive tasks are known to be sexually dimorphic. The better verbal ability of women is associated with reduced lateralization of language tasks while the male advantage in spatial tasks corresponds to marked cerebral lateralization. Sexual orientation effects in some of these tasks have been found in recent studies.
In the Vincent Mechanical Diagrams test, a dot detection divided field measure of functional cerebral asymmetry, homosexual men performed the same as heterosexual women with both scoring lower than heterosexual men displaying less asymmetry. Additionally, homosexual men display higher verbal performance IQ scores on subtests of the Wechsler Adult Intelligence Scale, in concordance with female testing patterns.[64] On several other tests, including a male-biased targeted throwing task and a female-biased Purdue Pegboard Test, the performance of homosexual men and heterosexual women showed no statistical difference from each other, while both significantly differed from heterosexual men.
Additionally, reduced asymmetry was found in a magnetoencephelographic study in which MEG-based source location estimates of an auditory evoked signal is found to be hemispherically symmetric in heterosexual women and homosexual men, while asymmetric in heterosexual men.[64]
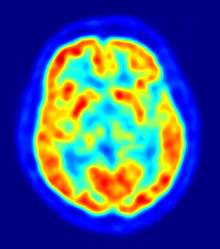
Response to serotonin
Serotonin is a neurotransmitter found in the central nervous system that has various roles in the regulation of sexual behavior. Serotonin agonists and antagonists have activational or inhibitory effects depending on their concentration and the brain area involved. Fluoxetine is a selective serotonin reuptake inhibitor that prolongs the effect of serotonin on neurons.[65] Kinnunen et al. (2004) administered fluoxetine to their study subjects to see if the brain is differentially activated in homosexual and heterosexual men through the action of serotonin.[65][66] After fluorexine administration, they measured glucose metabolism in the brain by using fluorodeoxyglucose positron emission tomography (FDG-PET). They found that the brain response to fluoxetine differs between homosexual and heterosexual men, that is, homosexual men show a smaller reduction of glucose metabolism in the hypothalamus than heterosexual men. In addition, other areas of the brain were differentially activated: the prefrontal association cortex of homosexual men exhibited increased activity after fluoxetine administration while the prefrontal association cortex of heterosexual men did not show any change. The cunate gyrus, lateral anterior cingulate and bilateral hippocampus/ parahippocampal gyrus of heterosexual men exhibited increased activity while decreased activity was seen in portions of their cingulate cortex.[66] These findings suggest that homosexuals and heterosexuals may not only differ in the total number of neurons in various areas of their central nervous systems, but also may differ in the distribution of certain kinds of neurons, such as serotonergic and dopaminergic neurons.[65]
Related studies
Various animal and insect models have been used to explore sexual orientation and brain characteristics. One experiment involved genetically altering male Drosophila causing them to have feminized brain structures involved in processing sexually dimorphic contact pheromones. Transformed males showed increase homosexual courtship behaviors to wild-type male flies, and there a correlation was found between the courtship behavior and the expression of the altered gene in the sexually related brain regions.[67]
Future studies
The development of sexual orientation is a far from complete subject. While neuroscience has made advancements shedding light on the mechanisms and relationships between the human brain and sexual orientation, much more further research should be conducted.
Areas for future research include:[3]
- finding markers for sex steroid levels in the brains of fetuses that highlight features of early neuro-development leading to certain sexual orientations
- determine the precise neural circuitry underlying direction of sexual preference
- use animal models to explore genetic and developmental factors that influence sexual orientation
- further population studies, genetic studies, and serological markers to clarify and definitively determine the effect of maternal immunity
- neuroimaging studies to quantify sexual-orientation-related differences in structure and function in vivo
- neurochemical studies to investigate the roles of sex steroids upon neural circuitry involved in sexual attraction
Notes
- ↑ The term nucleus in neuroanatomy must not be confused with the term nucleus in cytology. In cytology, the term nucleus refers to the organelle found in eukaryotic cells that contains the cell's genetic material. In neuroanatomy, the term nucleus refers to discrete groups of densely packed neuronal cell bodies in the central nervous system.[38][39] In anatomical sections, a nucleus appears as a region of gray matter surrounded by white matter.
- ↑ Another study showed that the cortical thickness of the right pars triangularis also differs between MtF transsexuals and homosexual men. Specifically, the pars triangularis of MtF transsexuals (and heterosexual men) is thicker than the pars triangularis of homosexual men. Furthermore, the pars triangularis of MtF transsexuals is thicker than the pars triangularis of heterosexual men. Notably, in both studies, the affected region is the right pars triangularis (i.e., the pars triangularis in the right hemisphere).[56]
References
- ↑ "Sexual orientation, homosexuality and bisexuality". American Psychological Association. Archived from the original on August 8, 2013. Retrieved August 10, 2013.
- ↑ "Sexual Orientation". American Psychiatric Association. Archived from the original on July 22, 2011. Retrieved January 1, 2013.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Rahman, Q (2005). "The neurodevelopment of human sexual orientation". Neuroscience & Biobehavioral Reviews. 29 (7): 1057–66. doi:10.1016/j.neubiorev.2005.03.002. PMID 16143171.
- ↑ Williams TJ; Pepitone ME; Christensen SE; Cooke BM; Huberman AD; Breedlove NJ; Breedlove TJ; Jordan CL; Breedlove SM (Mar 2000). "Finger-length ratios and sexual orientation" (PDF). Nature. 404 (6777): 455–6. doi:10.1038/35006555. PMID 10761903. Retrieved 17 August 2016.
- ↑ Swaab DF (December 2004). "Sexual differentiation of the human brain: relevance for gender identity, transsexualism and sexual orientation". Gynecological Endocrinology. 19 (6): 301–12. doi:10.1080/09513590400018231. PMID 15724806.
- ↑ Brown et al. 2002
- ↑ Okten et al. 2002
- ↑ Berenbaum SA, Bryk KK, Nowak N, Quigley CA, Moffat S (November 2009). "Fingers as a marker of prenatal androgen exposure". Endocrinology. 150 (11): 5119–24. doi:10.1210/en.2009-0774. PMC 2775980
 . PMID 19819951.
. PMID 19819951. - ↑ Manning, John T.; Bundred, Peter E.; Newton, Darren J.; Flanagan, Brian F. (2003). "The second to fourth digit ratio and variation in the androgen receptor gene". Evolution and Human Behavior. 24 (6): 399–405. doi:10.1016/S1090-5138(03)00052-7.
- ↑ Williams, T. J.; Pepitone, ME; Christensen, SE; Cooke, BM; Huberman, AD; Breedlove, NJ; Breedlove, TJ; Jordan, CL; Breedlove, SM (March 2000). "Finger-length ratios and sexual orientation" (PDF). Nature. 404 (6777): 455–456. doi:10.1038/35006555. PMID 10761903.
- ↑ Tortorice, J.L. (2002). "Written on the body: butch/femme lesbian gender identity and biological correlates". Rutgers Ph.D. Dissertation.
- ↑ McFadden D, Shubel E (December 2002). "Relative lengths of fingers and toes in human males and females". Hormones and Behavior. 42 (4): 492–500. doi:10.1006/hbeh.2002.1833. PMID 12488115.
- ↑ Hall LS, Love CT (February 2003). "Finger-length ratios in female monozygotic twins discordant for sexual orientation". Archives of Sexual Behavior. 32 (1): 23–8. doi:10.1023/A:1021837211630. PMID 12597269.
- 1 2 3 Rahman Q; Wilson GD (Apr 2003). "Sexual orientation and the 2nd to 4th finger length ratio: evidence for organising effects of sex hormones or developmental instability?". Psychoneuroendocrinology. Elsevier Inc. 28 (3): 288–303. doi:10.1016/S0306-4530(02)00022-7. PMID 12573297.
- ↑ Csathó A, Osváth A, Bicsák E, Karádi K, Manning J, Kállai J (February 2003). "Sex role identity related to the ratio of second to fourth digit length in women". Biological Psychology. 62 (2): 147–56. doi:10.1016/S0301-0511(02)00127-8. PMID 12581689.
- ↑ Putz, D; Gaulin, Steven J.C.; Sporter, Robert J.; McBurney, Donald H. (2004). "Sex hormones and finger lengthWhat does 2D:4D indicate?" (PDF). Evolution and Human Behavior. 25 (3): 182. doi:10.1016/j.evolhumbehav.2004.03.005.
- ↑ Rahman Q (May 2005). "Fluctuating asymmetry, second to fourth finger length ratios and human sexual orientation". Psychoneuroendocrinology. 30 (4): 382–91. doi:10.1016/j.psyneuen.2004.10.006. PMID 15694118.
- ↑ Kraemer B, Noll T, Delsignore A, Milos G, Schnyder U, Hepp U (2006). "Finger length ratio (2D:4D) and dimensions of sexual orientation". Neuropsychobiology. 53 (4): 210–4. doi:10.1159/000094730. PMID 16874008.
- ↑ Wallien MS, Zucker KJ, Steensma TD, Cohen-Kettenis PT (August 2008). "2D:4D finger-length ratios in children and adults with gender identity disorder". Hormones and Behavior. 54 (3): 450–4. doi:10.1016/j.yhbeh.2008.05.002. PMID 18585715.
- ↑ Bogaert AF (July 2006). "Biological versus nonbiological older brothers and men's sexual orientation". Proceedings of the National Academy of Sciences of the United States of America. 103 (28): 10771–4. doi:10.1073/pnas.0511152103. PMC 1502306. PMID 16807297.
- 1 2 3 4 Bogaert AF; Skorska M (2011). "Sexual orientation, fraternal birth order, and the maternal immune hypothesis: a review". Front Neuroendocrinol. 32 (2): 247–54. doi:10.1016/j.yfrne.2011.02.004. PMID 21315103.
- ↑ Cantor JM, Blanchard R, Paterson AD, Bogaert AF (February 2002). "How many gay men owe their sexual orientation to fraternal birth order?". Arch Sex Behav. 31 (1): 63–71. doi:10.1023/A:1014031201935. PMID 11910793.
- ↑ Blanchard R, Bogaert AF (January 1996). "Homosexuality in men and number of older brothers". Am J Psychiatry. 153 (1): 27–31. doi:10.1176/ajp.153.1.27. PMID 8540587.
- ↑ Blanchard R; Bogaert AF (2004). "Proportion of homosexual men who owe their sexual orientation to fraternal birth order: An estimate based on two national probability samples.". Am J Hum Biol. 16 (2): 151–7. doi:10.1002/ajhb.20006. PMID 14994314.
- 1 2 Blanchard R, Lippa RA (April 2007). "Birth order, sibling sex ratio, handedness, and sexual orientation of male and female participants in a BBC internet research project". Archives of Sexual Behavior. 36 (2): 163–76. doi:10.1007/s10508-006-9159-7. PMID 17345165.
- 1 2 3 4 5 6 7 Rahman Q; Wilson GD (2003). "Born gay? The psychobiology of human sexual orientation". Personality and Individual Differences. Elsevier. 34: 1337–1382.
- 1 2 3 Lalumière ML, Blanchard R, Zucker KJ (July 2000). "Sexual orientation and handedness in men and women: a meta-analysis". Psychological Bulletin. 126 (4): 575–92. doi:10.1037/0033-2909.126.4.575. PMID 10900997.
- ↑ Rahman Q (May 2005). "Fluctuating asymmetry, 2nd to 4th finger length ratios and human sexual orientation.". Psychoneuroendocrinology. Elsevier Inc. 30 (4): 382–91. doi:10.1016/j.psyneuen.2004.10.006. PMID 15694118.
- ↑ Mustanski BS; Bailey JM; Kaspar S (Feb 2002). "Dermatoglyphics, handedness, sex, and sexual orientation.". Arch Sex Behav. 31 (1): 113–22. doi:10.1023/A:1014039403752. PMID 11910784.
- 1 2 3 Simon LeVay (2010). Gay, Straight, and the Reason Why: The Science of Sexual Orientation. Oxford University Press. pp. 230–5. ISBN 0199752966.
- ↑ Martin JT; Puts DA; Breedlove SM (Mar 2008). "Hand Asymmetry in Heterosexual and Homosexual Men and Women: Relationship to 2D:4D Digit Ratios and Other Sexually Dimorphic Anatomical Traits". Arch Sex Behav. 37: 119–132. doi:10.1007/s10508-007-9279-8. PMID 18161017.
- ↑ Innala SM; Ernulf KE (Jun 1994). "When gay is pretty: physical attractiveness and low homophobia". Psychol Rep. 74 (3 Pt 1): 827–31. PMID 8058859.
- ↑ Minna Lyons; et al. (2014). "Detection of Sexual Orientation (Gaydar) by Homosexual and Heterosexual Women". Arch Sex Behav. 43: 345–352. doi:10.1007/s10508-013-0144-7.
- ↑ Zucker KJ; Wild J; Bradley SJ; Lowry CB (Feb 1993). "Physical attractiveness of boys with gender identity disorder". Arch Sex Behav. Kluwer Academic Publishers-Plenum Publishers. 22 (1): 23–36. doi:10.1007/BF01552910. ISSN 1573-2800. PMID 8435037.
- ↑ Rhodes G; Hickford C; Jeffery L (Feb 2000). "Sex-typicality and attractiveness: are supermale and superfemale faces super-attractive?". Br J Psychol. 91 (1): 125–40. doi:10.1348/000712600161718. PMID 10717775.
- ↑ Thornhill R; Gangestad SW (Dec 1999). "Facial attractiveness". Trends Cogn Sci. 3 (12): 452–460. doi:10.1016/S1364-6613(99)01403-5. PMID 10562724.
- ↑ Gangestad SW; Bailey JM; Martin NG (Jun 2000). "Taxometric analyses of sexual orientation and gender identity". J. Pers. Soc. Psychol. 78 (6): 1109–21. doi:10.1037/0022-3514.78.6.1109. PMID 10870912.
- ↑ Snell, Richard S (2010). Clinical Neuroanatomy (7th ed.). Lippincott Williams & Wilkins. p. 34. ISBN 9780781759939.
- ↑ Blumenfeld, Hal (2010). Neuroanatomy through clinical cases (2nd ed.). Sunderland, Mass.: Sinauer Associates. p. 21. ISBN 9780878936137.
- 1 2 3 4 5 6 Swaab DF, Hofman MA (June 1995). "Sexual differentiation of the human hypothalamus in relation to gender and sexual orientation". Trends in Neurosciences. 18 (6): 264–70. doi:10.1016/0166-2236(95)80007-O. PMID 7571001.
- ↑ LeVay S (August 1991). "A difference in hypothalamic structure between heterosexual and homosexual men". Science. 253 (5023): 1034–7. doi:10.1126/science.1887219. PMID 1887219.
- 1 2 Byne W; Lasco MS; Kemether E; Shinwari A; Edgar MA; Morgello S; Jones LB; Tobet S. (21 Feb 2000). "The interstitial nuclei of the human anterior hypothalamus: an investigation of sexual variation in volume and cell size, number and density". Brain Res. 856 (1-2): 254–8. doi:10.1016/S0006-8993(99)02458-0. ISSN 0006-8993. PMID 10677635.
- ↑ Swaab DF; Hofman MA. (24 Dec 1990). "An enlarged suprachiasmatic nucleus in homosexual men.". Brain Res. 537 (1-2): 141–8. doi:10.1016/0006-8993(90)90350-K. PMID 2085769.
- 1 2 3 4 Abe C; Johansson E; Allzen E; Savic I (Dec 2014). Gasbarri, Antonella, ed. "Sexual Orientation Related Differences in Cortical Thickness in Male Individuals". PLoS ONE. 9 (12): e114721. doi:10.1371/journal.pone.0114721. PMID 25479554.
- ↑ Hu S; Xu D; Peterson B; Wang Q; He X; et al. (2013). "Association of cerebral networks in resting state with sexual preference of homosexual men: a study of regional homogeneity and functional connectivity". PloS one. 8: e59426. doi:10.1371/journal.pone.0059426. PMID 23555670.
- ↑ Aste N; Balthazart J; Absil P; Grossmann R; Mülhbauer E; Viglietti-Panzica C; Panzica GC (29 Jun 1998). "Anatomical and neurochemical definition of the nucleus of the stria terminalis in Japanese quail (Coturnix japonica).". J Comp Neurol. 396 (2): 141–57. doi:10.1002/(SICI)1096-9861(19980629)396:2<141::AID-CNE1>3.0.CO;2-0. PMID 9634138.
- 1 2 Aldo Poiani (Aug 2010). Animal Homosexuality: A Biosocial Perspective. Cambridge University Press. p. 213. ISBN 9781139490382.
- 1 2 Zhou, JN; Hofman, MA; Gooren, LJG; Swaab, DF (Nov 1995). "A sex difference in the human brain and its relation to transsexuality". Nature. 378 (6552): 68–70. doi:10.1038/378068a0. PMID 7477289.
- 1 2 3 Savic I, Lindström P (July 2008). "PET and MRI show differences in cerebral asymmetry and functional connectivity between homo- and heterosexual subjects". Proceedings of the National Academy of Sciences of the United States of America. 105 (27): 9403–8. doi:10.1073/pnas.0801566105. PMC 2453705. PMID 18559854.
- 1 2 3 Hill AK; Dawood K; Puts DA (2012). "Biological Foundations of Sexual Orientation". Handbook of Psychology and Sexual Orientation (illustrated ed.). OUP USA, 2013. pp. 55–68. ISBN 9780199765218.
- 1 2 3 4 Witelson SF; Kigar DL; Scamvougeras A; Kideckel DM; Buck B; Stanchev PL; Bronskill M; Black S (Dec 2008). "Corpus callosum anatomy in right-handed homosexual and heterosexual men". Arch Sex Behav. 37 (6): 857–63. doi:10.1007/s10508-007-9276-y. ISSN 1573-2800. PMID 17975723.
- 1 2 Aldo Poiani (Aug 2010). Animal Homosexuality: A Biosocial Perspective. Cambridge University Press. pp. 213–5. ISBN 9781139490382.
- ↑ Simon LeVay (2010). Gay, Straight, and the Reason Why: The Science of Sexual Orientation. Oxford University Press, 2010. pp. 201–5. ISBN 0199752966.
- 1 2 3 4 5 Ponseti J; Siebner HR; Klöppel S; Wolff S; Granert O; Jansen O; Mehdorn HM; Bosinski HA (22 Aug 2007). "Homosexual Women Have Less Grey Matter in Perirhinal Cortex than Heterosexual Women". PLoS ONE. 2 (8): e762. doi:10.1371/journal.pone.0000762. PMC 1942120.
- ↑ Aldo Poiani (Aug 2010). Animal Homosexuality: A Biosocial Perspective. Cambridge University Press. p. 220. ISBN 9781139490382.
- ↑ Guillamon A; Junque C; Gómez-Gil E (Oct 2016). "A Review of the Status of Brain Structure Research in Transsexualism". Arch Sex Behav. Springer US. 45 (7): 1615–1648. doi:10.1007/s10508-016-0768-5. ISSN 1573-2800. PMID 27255307.
- 1 2 Berglund H, Lindström P, Savic I (May 2006). "Brain response to putative pheromones in lesbian women". Proceedings of the National Academy of Sciences of the United States of America. 103 (21): 8269–74. doi:10.1073/pnas.0600331103. PMC 1570103. PMID 16705035.
- 1 2 Savic I, Berglund H, Lindström P (May 2005). "Brain response to putative pheromones in homosexual men". Proceedings of the National Academy of Sciences of the United States of America. 102 (20): 7356–61. doi:10.1073/pnas.0407998102. PMC 1129091. PMID 15883379.
- 1 2 Safron A; Barch B; Bailey JM; Gitelman DR; Parrish TB; Reber PJ (Apr 2007). "Neural correlates of sexual arousal in homosexual and heterosexual men.". Behav Neurosci. 121 (2): 237–48. doi:10.1037/0735-7044.121.2.237. PMID 17469913.
- ↑ Paul T, Schiffer B, Zwarg T, et al. (June 2008). "Brain response to visual sexual stimuli in heterosexual and homosexual males". Human Brain Mapping. 29 (6): 726–35. doi:10.1002/hbm.20435. PMID 17636559.
- ↑ Hu SH; Wei N; Wang QD; et al. (Nov 2008). "Patterns of brain activation during visually evoked sexual arousal differ between homosexual and heterosexual men.". AJNR Am J Neuroradiol. 29 (10): 1890–6. doi:10.3174/ajnr.A1260. PMID 18768725.
- ↑ Ponseti J; Bosinski HA; Wolff S; Peller M; Jansen O; Mehdorn HM; Büchel C; Siebner HR (15 Nov 2006). "A functional endophenotype for sexual orientation in humans.". Neuroimage. 33 (3): 825–33. doi:10.1016/j.neuroimage.2006.08.002. PMID 16979350.
- ↑ Kranz F; Ishai A (2006). "Face Perception Is Modulated by Sexual Preference". Current Biology. 16 (1): 63–68. doi:10.1016/j.cub.2005.10.070.
- 1 2 3 Sanders G, Wright M (October 1997). "Sexual orientation differences in cerebral asymmetry and in the performance of sexually dimorphic cognitive and motor tasks". Archives of Sexual Behavior. 26 (5): 463–80. doi:10.1023/A:1024551704723. PMID 9343633.
- 1 2 3 Aldo Poiani (Aug 2010). Animal Homosexuality: A Biosocial Perspective (illustrated ed.). Cambridge University Press. pp. 222–4. ISBN 9781139490382.
- 1 2 Kinnunen LH; Moltz H; Metz J; Cooper M (22 Oct 2004). "Differential brain activation in exclusively homosexual and heterosexual men produced by the selective serotonin reuptake inhibitor, fluoxetine". Brain Res. 1024 (1-2): 251–4. doi:10.1016/j.brainres.2004.07.070. PMID 15451388.
- ↑ Ferveur JF, Störtkuhl KF, Stocker RF, Greenspan RJ (February 1995). "Genetic feminization of brain structures and changed sexual orientation in male Drosophila". Science. 267 (5199): 902–5. doi:10.1126/science.7846534. PMID 7846534.
| |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
