Mycobacterium tuberculosis
| Mycobacterium tuberculosis | |
|---|---|
 | |
| M. tuberculosis bacterial colonies | |
| Scientific classification | |
| Domain: | Bacteria |
| Phylum: | Actinobacteria |
| Class: | Actinobacteria |
| Order: | Actinomycetales |
| Family: | Mycobacteriaceae |
| Genus: | Mycobacterium |
| Species: | M. tuberculosis |
| Binomial name | |
| Mycobacterium tuberculosis Zopf 1883 | |
| Synonyms | |
|
Tubercle bacillus Koch 1882 | |
Mycobacterium tuberculosis is an obligate[1] pathogenic bacterial species in the family Mycobacteriaceae and the causative agent of tuberculosis.[2] First discovered in 1882 by Robert Koch, M. tuberculosis has an unusual, waxy coating on its cell surface (primarily due to the presence of mycolic acid), which makes the cells impervious to Gram staining; M. tuberculosis can appear Gram-negative and Gram-positive in clinical settings.[3] The Lowenstein-Jensen Agar, or acid-fast stain, is used instead. The physiology of M. tuberculosis is highly aerobic and requires high levels of oxygen. Primarily a pathogen of the mammalian respiratory system, it infects the lungs. The most frequently used diagnostic methods for tuberculosis are the tuberculin skin test, acid-fast stain, and chest radiographs.[2]
The M. tuberculosis genome was sequenced in 1998.[4][5]
Pathophysiology
M. tuberculosis requires oxygen to grow and can be cultured in the laboratory.[6] It does not retain any bacteriological stain due to high lipid content in its wall, hence Ziehl-Neelsen staining, or acid-fast staining, is used. Mycobacteria have an outer membrane.[7]
M. tuberculosis divides every 15–20 hours, which is extremely slow compared with other bacteria, which tend to have division times measured in minutes (Escherichia coli can divide roughly every 20 minutes). It is a small bacillus that can withstand weak disinfectants and can survive in a dry state for weeks. Its unusual cell wall, rich in lipids (e.g., mycolic acid), is likely responsible for this resistance and is a key virulence factor.[8]
Humans are the only known reservoirs of M. tuberculosis. There are misconceptions that M. tuberculosis can be spread by shaking hands, making contact with toilet seats, sharing food or drink, sharing toothbrushes, or kissing. M. tuberculosis can only be spread through air droplets originating from a person either coughing, sneezing, speaking, or singing that has the disease.[9]
When in the lungs, M. tuberculosis is taken up by alveolar macrophages, but they are unable to digest and eradicate the bacterium. Its cell wall prevents the fusion of the phagosome with the lysosome, which contains a host of antimycobacterial factors.[10] Specifically, M. tuberculosis blocks the bridging molecule, early endosomal autoantigen 1 (EEA1); however, this blockade does not prevent fusion of vesicles filled with nutrients. Consequently, the bacteria multiply unchecked within the macrophage. The bacteria also carry the UreC gene, which prevents acidification of the phagosome.[11] In addition, production of the diterpene isotuberculosinol prevents maturation of the phagosome.[12] The bacteria also evade macrophage-killing by neutralizing reactive nitrogen intermediates.[13]
Protective granulomas are formed due to the production of cytokines and upregulation of proteins involved in recruitment. Granulotomatous lesions are important in both regulating the immune response and minimizing tissue damage. Moreover, T cells help maintain Mycobacterium within the granulomas.[14]
The ability to construct M. tuberculosis mutants and test individual gene products for specific functions has significantly advanced the understanding of the pathogenesis and virulence factors of M. tuberculosis. Many secreted and exported proteins are known to be important in pathogenesis.[15] Aerolysin is a virulence factor of the pathogenic bacterium Aeromonas hydrophila. Resistant strains of mycobacterium tuberculosis have developed resistance to more than one TB drug, due to mutations in their genes.
Strain variation
Typing of strains is useful in the investigation of tuberculosis outbreaks, because it gives the investigator evidence for-or-against transmission from person to person. Consider the situation where person A has tuberculosis and believes he acquired it from person B. If the bacteria isolated from each person belong to different types, then transmission from B to A is definitively disproved; however, if the bacteria are the same strain, then this supports (but does not definitively prove) the hypothesis that B infected A.
Until the early 2000s, M. tuberculosis strains were typed by pulsed field gel electrophoresis (PFGE).[16] This has now been superseded by variable numbers of tandem repeats (VNTR), which is technically easier to perform and allows better discrimination between strains. This method makes use of the presence of repeated DNA sequences within the M. tuberculosis genome.
Three generations of VNTR typing for M. tuberculosis are noted. The first scheme, called exact tandem repeat, used only five loci,[17] but the resolution afforded by these five loci was not as good as PFGE. The second scheme, called mycobacterial interspersed repetitive unit, had discrimination as good as PFGE.[18][19] The third generation (mycobacterial interspersed repetitive unit - 2) added a further nine loci to bring the total to 24. This provides a degree of resolution greater than PFGE and is currently the standard for typing M. tuberculosis.[20] However, with regard to archaeological remains, additional evidence may be required because of possible contamination from related soil bacteria.[21]
Antibiotic resistance in M. tuberculosis typically occurs due to either the accumulation of mutations in the genes targeted by the antibiotic or a change in titration of the drug.[22] M. tuberculosis is considered to be multidrug-resistant (MDR TB) if it has developed drug resistance to both rifampicin and isoniazid, which are the most important antibiotics used in treatment. Additionally, extensively drug-resistant M. tuberculosis (XDR TB) is characterized by resistance to both isoniazid and rifampin, plus any fluoroquinolone and at least one of three injectable second-line drugs (i.e., amikacin, kanamycin, or capreomycin).[23]

Microscopy
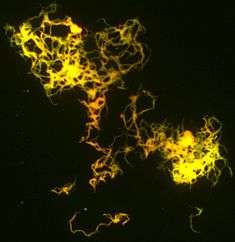
M. tuberculosis is characterized by caseating granulomas containing Langhans giant cells, which have a "horseshoe" pattern of nuclei. Organisms are identified by their red color on acid-fast staining. Cells are often seen wrapped together, due to the presence of fatty acids in the cell wall that stick together. This appearance is referred to as cording, like strands of cord that make up a rope.
Genome
The genome of the H37Rv strain was published in 1998.[24] Its size is 4 million base pairs, with 3959 genes; 40% of these genes have had their function characterised, with possible function postulated for another 44%. Within the genome are also six pseudogenes.
The genome contains 250 genes involved in fatty acid metabolism, with 39 of these involved in the polyketide metabolism generating the waxy coat. Such large numbers of conserved genes show the evolutionary importance of the waxy coat to pathogen survival. Furthermore, experimental studies have since validated the importance of a lipid metabolism for M. tuberculosis, consisting entirely of host-derived lipids such as fats and cholesterol. Bacteria isolated from the lungs of infected mice were shown to preferentially use fatty acids over carbohydrate substrates.[25] M. tuberculosis can also grow on the lipid cholesterol as a sole source of carbon, and genes involved in the cholesterol use pathway(s) have been validated as important during various stages of the infection lifecycle of M. tuberculosis, especially during the chronic phase of infection when other nutrients are likely not available.[26]
About 10% of the coding capacity is taken up by the PE/PPE gene families that encode acidic, glycine-rich proteins. These proteins have a conserved N-terminal motif, deletion of which impairs growth in macrophages and granulomas.[27]
Nine noncoding sRNAs have been characterised in M. tuberculosis,[28] with a further 56 predicted in a bioinformatics screen.[29]
In 2013, a study on the genome of several sensitive, ultraresistant, and multiresistant M. tuberculosis strains was made to study antibiotic resistance mechanisms. Results reveal new relationships and drug resistance genes not previously associated and suggest some genes and intergenic regions associated with drug resistance may be involved in the resistance to more than one drug. Noteworthy is the role of the intergenic regions in the development of this resistance, and most of the genes proposed in this study to be responsible for drug resistance have an essential role in the development of M. tuberculosis.[30]
Evolution
The M. tuberculosis complex evolved in Africa and most probably in the Horn of Africa.[31][32] The M. tuberculosis group has a number of members that include Mycobacterium africanum, Mycobacterium bovis (Dassie's bacillus), Mycobacterium caprae, Mycobacterium microti, Mycobacterium mungi, Mycobacterium orygis, and Mycobacterium pinnipedii. This group may also include the Mycobacterium canettii clade.
The M. canettii clade — which includes Mycobacterium prototuberculosis — is a group of smooth-colony Mycobacterium species. Unlike the established members of the M. tuberculosis group, they undergo recombination with other species. The majority of the known strains of this group have been isolated from the Horn of Africa. The ancestor of Mycobacterium tuberculosis appears to be the species Mycobacterium canettii, first described in 1969.[33]
The established members of the M. tuberculosis complex are all clonal in their spread. The main human-infecting species have been classified into seven spoligotypes: type 1 contains the East African-Indian (EAI) and some Manu (Indian) strains; type 2 is the Beijing group; type 3 consists of the Central Asian (CAS) strains; type 4 of the Ghana and Haarlem (H/T), Latin America-Mediterranean (LAM) and X strains; types 5 and 6 correspond to Mycobacterium africanum and are observed predominantly and at very high frequency in West Africa. A seventh type has been isolated from the Horn of Africa.[31] The other species of this complex belong to a number of spoligotypes and do not normally infect humans.
Types 2 and 3 are more closely related to each other than to the other types. Types 5 and 6 are most closely aligned with the species that do not normally infect humans. Type 3 has been divided into two clades: CAS-Kili (found in Tanzania) and CAS-Delhi (found in India and Saudi Arabia).
The most recent common ancestor of the M. tuberculosis complex evolved ~40,000 years ago.[34] The most recent common ancestor of the EAI and LAM strains has been estimated to be 13,700 and 7,000 years ago, respectively. The Beijing- CAS strains diverged about 17,100 years ago. All types of the M. tuberculosis began their current expansion about 5000 years ago—a period that coincides with the appearance of M. bovis. The Beijing strain appears to have been the most successful with around a 500-fold increase in effective population size (Ne) since its expansion began. The least successful of the main lineages appears to have been those limited to Africa, where they have undergone an Ne increase of only five-fold. Since its initial evolution, M. bovis has undergone an expansion of Ne of about 65-fold.
Co-evolution with modern humans
There is much evidence to suggest that the different strains of the obligate human pathogen M. tuberculosis have co-evolved, migrated, and expanded with their human hosts. This well-supported theory is consistent with the bacterium’s phylogeny and phylogeography.[32][35] With the global spread of M. tuberculosis, studies have examined whether geographically defined human populations are especially susceptible to the transmission of a particular lineage or strain of M. tuberculosis. They have found that even when transmission of M. tuberculosis occurs in an urban center outside the region of origin, a human host’s region of origin is predictive of which TB strain they carry and that genetically differentiated populations of M. tuberculosis do indeed preserve stable associations with host populations from their geographic region.[36][37] The fact that all six principle phylogeographic lineages are found in Africa combined with the belief that ancestral Mycobacteria may have impacted early hominids in East Africa as early as three million years ago, once again point to the theory of M. tuberculosis originating in Africa and expanding alongside the human migration out of East Africa.[38] The significant correlation of increased frequency of tuberculosis resistant alleles with the duration of a human population’s urban settlement similarly points to an extensive co-evolutionary relationship.[39] Some of the most compelling data concerning the co-expansion of M. tuberculosis with modern humans comes from a study that compared M. tuberculosis phylogeny to human mitochondrial genomes and found impressive similarities in the patterns and geographical locations of branching and divergence events.[32] The match between M. tuberculosis and human mitochondrial phylogenies supports an extended relationship between M. tuberculosis and its host, while the clear expansion of this bacterial pathogen during the Neolithic Demographic Transition (~10 000 years ago) suggests that M. tuberculosis was able to adapt to changing human populations and that the historical success of this pathogen was driven at least in part by dramatic increases in human host population density.
Host genetics
The nature of the host-pathogen interaction between humans and Mycobacterium tuberculosis is considered to have a genetic component. A group of rare disorders called Mendelian Susceptibility to Mycobacterial Diseases (MSMD) was observed in a subset of individuals with a genetic defect that results in increased susceptibility to Mycobacterial infection.
Early case and twin studies have indicated that genetic component are important in host susceptibility to Mycobacterial tuberculosis. Recent Genome-wide association studies (GWAS) have identified three genetic risk loci, including at positions 11p13 and 18q11.[40][41] As is common in GWAS, the variants discovered have moderate effect sizes.
DNA repair
As an intracellular pathogen M. tuberculosis is exposed to a variety of DNA damaging assaults, primarily from host-generated antimicrobial toxic radicals. Exposure to reactive oxygen species and/or reactive nitrogen species causes different types of DNA damage including oxidation, depurination, methylation and deamination that can give rise to single- and double-strand breaks (DSBs).
DnaE2 polymerase is upregulated in M. tuberculosis by several DNA damaging agents as well as during infection of mice.[42] Loss of this DNA polymerase reduces the virulence of M. tuberculosis in mice.[42] DnaE2 is an error-prone repair DNA repair polymerase that appears to contribute to M. tuberculosis survival during infection.
The two major pathways employed in repair of DSBs are homologous recombinational repair (HR) and non-homologous end joining (NHEJ). Macrophage-internalized M. tuberculosis is able to persist if either of these pathways is defective, but is attenuated when both pathways are defective.[43] This indicates that intracellular exposure of M. tuberculosis to reactive oxygen and/or reactive nitrogen species results in the formation of DSBs that are repaired by HR or NHEJ.[43] However deficiency of DSB repair does not appear to impair M. tuberculosis virulence in animal models.[44]
History
M. tuberculosis, then known as the "tubercle bacillus", was first described on 24 March 1882 by Robert Koch, who subsequently received the Nobel Prize in physiology or medicine for this discovery in 1905; the bacterium is also known as "Koch's bacillus".[45]
Tuberculosis has existed throughout history, but the name has changed frequently over time. In 1720, though, the history of tuberculosis started to take shape into what is known of it today; as the physician Benjamin Marten described in his A Theory of Consumption, tuberculosis may be caused by small living creatures transmitted through the air to other patients.[46]
Vaccine
The BCG vaccine has had success in preventing tuberculosis.
See also
References
- ↑ Asunción Martínez; Sandra Torello; Roberto Kolter (1999). "Sliding Motility in Mycobacteria". J Bacteriol. 181 (23): 7331–7338. PMC 103697
 . PMID 10572138.
. PMID 10572138. - 1 2 Ryan, Kenneth J.; Ray, C. George (2004). "Mycobacteria". Sherris Medical Microbiology : an Introduction to Infectious Diseases (4th ed.). New York: McGraw-Hill. p. 439. ISBN 0-83-858529-9.
- ↑ Fu, L. M.; Fu-Liu, C. S. (2002-01-01). "Is Mycobacterium tuberculosis a closer relative to Gram-positive or Gram-negative bacterial pathogens?". Tuberculosis (Edinburgh, Scotland). 82 (2-3): 85–90. doi:10.1054/tube.2002.0328. ISSN 1472-9792. PMID 12356459.
- ↑ Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C, Harris D, Gordon SV, Eiglmeier K, Gas S, Barry CE, Tekaia F, Badcock K, Basham D, Brown D, Chillingworth T, Connor R, Davies R, Devlin K, Feltwell T, Gentles S, Hamlin N, Holroyd S, Hornsby T, Jagels K, Krogh A, McLean J, Moule S, Murphy L, Oliver K, Osborne J, Quail MA, Rajandream MA, Rogers J, Rutter S, Seeger K, Skelton J, Squares R, Squares S, Sulston JE, Taylor K, Whitehead S, Barrell BG (June 1998). "Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence". Nature. 393 (6685): 537–44. doi:10.1038/31159. PMID 9634230.
- ↑ Camus JC, Pryor MJ, Médigue C, Cole ST (October 2002). "Re-annotation of the genome sequence of Mycobacterium tuberculosis H37Rv". Microbiology (Reading, Engl.). 148 (Pt 10): 2967–73. doi:10.1099/00221287-148-10-2967. PMID 12368430.
- ↑ Parish T, Stoker NG (1999). "Mycobacteria: bugs and bugbears (two steps forward and one step back)". Mol. Biotechnol. 13 (3): 191–200. doi:10.1385/MB:13:3:191. PMID 10934532.
- ↑ Niederweis M, Danilchanka O, Huff J, Hoffmann C, Engelhardt H (2010). "Mycobacterial outer membranes: in search of proteins". Trends in Microbiology. 18 (3): 109–16. doi:10.1016/j.tim.2009.12.005. PMC 2931330. PMID 20060722.
- ↑ Murray PR, Rosenthal KS, Pfaller MA (2005). Medical Microbiology. Elsevier Mosby.
- ↑ Center for Disease Control (April 2016). "Basic TB Facts".
- ↑ Keane J, Balcewicz-Sablinska MK, Remold HG, Chupp GL, Meek BB, Fenton MJ, Kornfeld H (1997). "Infection by Mycobacterium tuberculosis promotes human alveolar macrophage apoptosis". Infect. Immun. 65 (1): 298–304. PMC 174591. PMID 8975927.
- ↑ Bell E (October 2005). "Vaccines: A souped-up version of BCG". Nature Reviews Immunology. 5 (10): 746. doi:10.1038/nri1720.
- ↑ Mann FM, Xu M, Chen X, Fulton DB, Russell DG, Peters RJ (2009). "Edaxadiene: A New Bioactive Diterpene fromMycobacterium tuberculosis". Journal of the American Chemical Society. 131 (48): 17526–17527. doi:10.1021/ja9019287. PMC 2787244. PMID 19583202.
- ↑ Flynn JL, Chan J (August 2003). "Immune evasion by Mycobacterium tuberculosis: living with the enemy". Curr. Opin. Immunol. 15 (4): 450–5. doi:10.1016/S0952-7915(03)00075-X. PMID 12900278.
- ↑ Saunders B. M.; Cooper A. M. (2000). "Restraining mycobacteria: role of granulomas in mycobacterial infections". Immunology and cell biology. 78 (4): 334–341. doi:10.1046/j.1440-1711.2000.00933.x.
- ↑ Wooldridge K, ed. (2009). Bacterial Secreted Proteins: Secretory Mechanisms and Role in Pathogenesis. Caister Academic Press. ISBN 978-1-904455-42-4.
- ↑ Zhang Y, Mazurek GH, Cave MD, Eisenach KD, Pang Y, Murphy DT, Wallace RJ (June 1992). "DNA polymorphisms in strains of Mycobacterium tuberculosis analyzed by pulsed-field gel electrophoresis: a tool for epidemiology". J. Clin. Microbiol. 30 (6): 1551–6. PMC 265327. PMID 1352518.
- ↑ Frothingham R, Meeker-O'Connell WA (1998). "Genetic diversity in the Mycobacterium tuberculosis complex based on variable numbers of tandem DNA repeats". Microbiology. 144 (Pt 5): 1189–96. doi:10.1099/00221287-144-5-1189. PMID 9611793.
- ↑ Mazars E, Lesjean S, Banuls AL, Gilbert M, Vincent V, Gicquel B, Tibayrenc M, Locht C, Supply P (2001). "High-resolution minisatellite-based typing as a portable approach to global analysis of Mycobacterium tuberculosis molecular epidemiology". Proc Natl Acad Sci U S A. 98 (4): 1901–6. doi:10.1073/pnas.98.4.1901. PMC 29354. PMID 11172048.
- ↑ Hawkey PM, Smith EG, Evans JT, Monk P, Bryan G, Mohamed HH, Bardhan M, Pugh RN (2003). "Mycobacterial interspersed repetitive unit typing of Mycobacterium tuberculosis compared to IS6110-based restriction fragment length polymorphism analysis for investigation of apparently clustered cases of tuberculosis". J Clin Microbiol. 41 (8): 3514–20. doi:10.1128/JCM.41.8.3514-3520.2003. PMC 179797. PMID 12904348.
- ↑ Supply P, Allix C, Lesjean S, Cardoso-Oelemann M, Rüsch-Gerdes S, Willery E, Savine E, de Haas P, van Deutekom H, Roring S, Bifani P, Kurepina N, Kreiswirth B, Sola C, Rastogi N, Vatin V, Gutierrez MC, Fauville M, Niemann S, Skuce R, Kremer K, Locht C, van Soolingen D (2006). "Proposal for standardization of optimized mycobacterial interspersed repetitive unit-variable-number tandem repeat typing of Mycobacterium tuberculosis". J Clin Microbiol. 44 (12): 4498–510. doi:10.1128/JCM.01392-06. PMC 1698431. PMID 17005759.
- ↑ Müller, Romy; Roberts, Charlotte A.; Brown, Terence A. (2015). "Complications in the study of ancient tuberculosis: non-specificity of IS6110 PCRs". Science and Technology of Archaeological Research. 1 (1): STAR20141120548. doi:10.1179/2054892314Y.0000000002.
- ↑ Rattan A, Kalia A, Ahmad N (June 1998). "Multidrug-Resistant Mycobacterium tuberculosis: Molecular Perspectives". Emerg Infect Dis. 4: 195–209. doi:10.3201/eid0402.980207.
- ↑ Center for Disease Control (April 2014). "Drug-resistant TB".
- ↑ "Mycobacterium tuberculosis". Sanger Institute. 2007-03-29. Retrieved 2008-11-16.
- ↑ Bloch H, Segal W (August 1956). "Biochemical differentiation of Mycobacterium tuberculosis grown in vivo and in vitro". J. Bacteriol. 72 (2): 132–41. PMC 357869. PMID 13366889.
- ↑ Wipperman MF, Sampson NS, Thomas ST (2014). "Pathogen roid rage: Cholesterol utilization by Mycobacterium tuberculosis". Crit. Rev. Biochem. Mol. Biol. 49 (4): 269–93. doi:10.3109/10409238.2014.895700. PMID 24611808.
- ↑ Glickman MS, Jacobs WR (February 2001). "Microbial pathogenesis of Mycobacterium tuberculosis: dawn of a discipline". Cell. 104 (4): 477–85. doi:10.1016/S0092-8674(01)00236-7. PMID 11239406.
- ↑ Arnvig KB, Young DB (August 2009). "Identification of small RNAs in Mycobacterium tuberculosis". Mol. Microbiol. 73 (3): 397–408. doi:10.1111/j.1365-2958.2009.06777.x. PMC 2764107. PMID 19555452.
- ↑ Livny J, Brencic A, Lory S, Waldor MK (2006). "Identification of 17 Pseudomonas aeruginosa sRNAs and prediction of sRNA-encoding genes in 10 diverse pathogens using the bioinformatic tool sRNAPredict2". Nucleic Acids Res. 34 (12): 3484–93. doi:10.1093/nar/gkl453. PMC 1524904. PMID 16870723.
- ↑ Zhang H, Li D, Zhao L, Fleming J, Lin N, Wang T, Liu Z, Li C, Galwey N, Deng J, Zhou Y, Zhu Y, Gao Y, Wang T, Wang S, Huang Y, Wang M, Zhong Q, Zhou L, Chen T, Zhou J, Yang R, Zhu G, Hang H, Zhang J, Li F, Wan K, Wang J, Zhang XE, Bi L (October 2013). "Genome sequencing of 161 Mycobacterium tuberculosis isolates from China identifies genes and intergenic regions associated with drug resistance". Nat. Genet. 45 (10): 1255–60. doi:10.1038/ng.2735. PMID 23995137.
- 1 2 Blouin Y, Hauck Y, Soler C, Fabre M, Vong R, Dehan C, Cazajous G, Massoure PL, Kraemer P, Jenkins A, Garnotel E, Pourcel C, Vergnaud G (2012). "Significance of the identification in the Horn of Africa of an exceptionally deep branching Mycobacterium tuberculosis clade". PLoS ONE. 7 (12): e52841. doi:10.1371/journal.pone.0052841. PMC 3531362. PMID 23300794.
- 1 2 3 Comas I, Coscolla M, Luo T, Borrell S, Holt KE, Kato-Maeda M, Parkhill J, Malla B, Berg S, Thwaites G, Yeboah-Manu D, Bothamley G, Mei J, Wei L, Bentley S, Harris SR, Niemann S, Diel R, Aseffa A, Gao Q, Young D, Gagneux S (2013). "Out-of-Africa migration and Neolithic coexpansion of Mycobacterium tuberculosis with modern humans". Nat. Genet. 45 (10): 1176–82. doi:10.1038/ng.2744. PMC 3800747. PMID 23995134.
- ↑ Blouin Y, Cazajous G, Dehan C, Soler C, Vong R, Hassan MO, Hauck Y, Boulais C, Andriamanantena D, Martinaud C, Martin É, Pourcel C, Vergnaud G (January 2014). "Progenitor Mycobacterium canettii clone responsible for lymph node tuberculosis epidemic, Djibouti". Emerging Infect. Dis. 20 (1): 21–8. doi:10.3201/eid2001.130652. PMC 3884719. PMID 24520560.
- ↑ Wirth T, Hildebrand F, Allix-Béguec C, Wölbeling F, Kubica T, Kremer K, van Soolingen D, Rüsch-Gerdes S, Locht C, Brisse S, Meyer A, Supply P, Niemann S (2008). "Origin, spread and demography of the Mycobacterium tuberculosis complex". PLoS Pathog. 4 (9): e1000160. doi:10.1371/journal.ppat.1000160. PMC 2528947. PMID 18802459.
- ↑ Hershberg R, Lipatov M, Small PM, Sheffer H, Niemann S, Homolka S, Roach JC, Kremer K, Petrov DA, Feldman MW, Gagneux S (2008). "High functional diversity in Mycobacterium tuberculosis driven by genetic drift and human demography". PLoS Biol. 6 (12): e311. doi:10.1371/journal.pbio.0060311. PMC 2602723. PMID 19090620.
- ↑ Gagneux S, DeRiemer K, Van T, Kato-Maeda M, de Jong BC, Narayanan S, Nicol M, Niemann S, Kremer K, Gutierrez MC, Hilty M, Hopewell PC, Small PM (2006). "Variable host-pathogen compatibility in Mycobacterium tuberculosis". Proc. Natl. Acad. Sci. U.S.A. 103 (8): 2869–73. doi:10.1073/pnas.0511240103. PMC 1413851. PMID 16477032.
- ↑ Hirsh AE, Tsolaki AG, DeRiemer K, Feldman MW, Small PM (2004). "Stable association between strains of Mycobacterium tuberculosis and their human host populations". Proc. Natl. Acad. Sci. U.S.A. 101 (14): 4871–6. doi:10.1073/pnas.0305627101. PMC 387341. PMID 15041743.
- ↑ Gutierrez MC, Brisse S, Brosch R, Fabre M, Omaïs B, Marmiesse M, Supply P, Vincent V (2005). "Ancient origin and gene mosaicism of the progenitor of Mycobacterium tuberculosis". PLoS Pathog. 1 (1): e5. doi:10.1371/journal.ppat.0010005. PMC 1238740. PMID 16201017.
- ↑ Barnes I, Duda A, Pybus OG, Thomas MG (2011). "Ancient urbanization predicts genetic resistance to tuberculosis". Evolution. 65 (3): 842–8. doi:10.1111/j.1558-5646.2010.01132.x. PMID 20840594.
- ↑ Thye T, Owusu-Dabo E, Vannberg FO, van Crevel R, Curtis J, Sahiratmadja E, Balabanova Y, Ehmen C, Muntau B, Ruge G, Sievertsen J, Gyapong J, Nikolayevskyy V, Hill PC, Sirugo G, Drobniewski F, van de Vosse E, Newport M, Alisjahbana B, Nejentsev S, Ottenhoff TH, Hill AV, Horstmann RD, Meyer CG (5 February 2012). "Common variants at 11p13 are associated with susceptibility to tuberculosis.". Nature Genetics. 44 (3): 257–9. doi:10.1038/ng.1080. PMID 22306650.
- ↑ Thye T, Vannberg FO, Wong SH, Owusu-Dabo E, Osei I, Gyapong J, Sirugo G, Sisay-Joof F, Enimil A, Chinbuah MA, Floyd S, Warndorff DK, Sichali L, Malema S, Crampin AC, Ngwira B, Teo YY, Small K, Rockett K, Kwiatkowski D, Fine PE, Hill PC, Newport M, Lienhardt C, Adegbola RA, Corrah T, Ziegler A, Morris AP, Meyer CG, Horstmann RD, Hill AV (September 2010). "Genome-wide association analyses identifies a susceptibility locus for tuberculosis on chromosome 18q11.2.". Nature Genetics. 42 (9): 739–41. doi:10.1038/ng.639. PMID 20694014.
- 1 2 Boshoff HI, Reed MB, Barry CE, Mizrahi V (2003). "DnaE2 polymerase contributes to in vivo survival and the emergence of drug resistance in Mycobacterium tuberculosis". Cell. 113 (2): 183–93. doi:10.1016/s0092-8674(03)00270-8. PMID 12705867.
- 1 2 Brzostek A, Szulc I, Klink M, Brzezinska M, Sulowska Z, Dziadek J (2014). "Either non-homologous ends joining or homologous recombination is required to repair double-strand breaks in the genome of macrophage-internalized Mycobacterium tuberculosis". PLoS ONE. 9 (3): e92799. doi:10.1371/journal.pone.0092799. PMC 3962454. PMID 24658131.
- ↑ Heaton BE, Barkan D, Bongiorno P, Karakousis PC, Glickman MS (2014). "Deficiency of double-strand DNA break repair does not impair Mycobacterium tuberculosis virulence in multiple animal models of infection". Infect. Immun. 82 (8): 3177–85. doi:10.1128/IAI.01540-14. PMC 4136208. PMID 24842925.
- ↑ "Robert Koch and Tuberculosis: Koch's Famous Lecture". Nobel Foundation. 2008. Retrieved 2008-11-18.
- ↑ "Tuberculosis History Timeline". Retrieved 2010-06-18.
External links
- TB database: an integrated platform for Tuberculosis research
- Photoblog about Tuberculosis
- "Mycobacterium tuberculosis". NCBI Taxonomy Browser.
- Database on Mycobacterium tuberculosis genetics
| Wikimedia Commons has media related to Mycobacterium tuberculosis. |