Lobopodia
| Lobopodia Fossil range: Cambrian Series 2–Early Pennsylvanian[1] The taxa Onychophora and Tardigrada survive to Recent | ||||
|---|---|---|---|---|
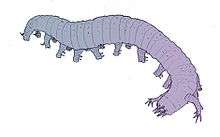 Reconstruction of the lobopod Aysheaia | ||||
| Scientific classification | ||||
| ||||
| Included groups | ||||
| ||||
| Excluded groups | ||||
|
Crown-group Euarthropoda | ||||
The lobopodians, members of the informal group Lobopodia Snodgrass 1938[2] (from the Greek, meaning "blunt feet") are worm-like taxa with stubby legs.
The scope of the Lobopodian concept varies from author to author. Its most limited sense refers to a suite of mainly Cambrian panarthropod taxa with flexible non-segmented limbs – for example Aysheaia, Hallucigenia and Xenusion. A broader definition of lobopodia would also incorporate the extant phyla Onychophora and Tardigrada. The broadest definition proposes the (monophyletic) Superphylum Lobopodia to encompass all three panarthropod phyla.[3]
The oldest near-complete fossil lobopodians date to the Lower Cambrian; some are also known from Ordovician, Silurian and Carboniferous Lagerstätten.[4][5][6] Some bear toughened claws, plates or spines, which are commonly preserved as carbonaceous or mineralized microfossils in Cambrian strata.[7][8]
Representative taxa
The better-known genera include, for example, Aysheaia, which was discovered in the Canadian Burgess Shale and looks something like the modern velvet worms., and Hallucigenia, known from Chengjiang and the Burgess Shale. The latter taxon was originally reconstructed with long, stilt-like legs and mysterious fleshy dorsal protuberances, and was long considered a prime example of the way in which nature experimented with the most diverse and bizarre body designs during the Cambrian.[9] However, further discoveries showed that this reconstruction had placed the animal upside-down: interpreting the "stilts" as dorsal spines made it clear that the fleshy "dorsal" protuberances were actually legs. This second reconstruction also exchanged the front and rear ends of the creature, which further investigation showed to be erroneous.[10] A study published in 2014 concluded that Hallucigenia is an ancestor of modern onychophora based on its claws.[11]
Microdictyon and Onychodictyon are charismatic genera resembling Hallucigenia, but with net-like plates instead of spines; members of these genera fall in the stem lineages of Onychophora and of Tardigrada.[11]
Other taxa such as Luolishania have elaborate fan-like feeding appendages that presumably formed 'baskets' for filter-feeding.[12]
Morphology
Most lobopodians are in the order of inches in length, though tardigrades are ~0.1 to 1.5 millimeters long. They are annulated, although the annulation may be difficult to discern, due to their close spacing (~0.2mm) and low relief.[13] Lobopodia and their legs are circular in cross-section.[13] Their legs, technically called lobopods, are loosely conical in shape, tapering from the body to their clawed tips.[13] The longest and most robust legs are at the middle of the trunk, with those nearer the head and tail more spindly.[13] The claws are slightly curved. Their length is loosely proportional to the length of the leg to which they are attached.[13] The eyes are similar to those of modern arthropods as has been shown in Miraluolishania haikouensis (Liu et al., 2004).
The gut of lobopodians is unusual in that it is straight, undifferentiated, and sometimes preserved in the fossil record in three dimensions. In some specimens the gut is found to be filled with sediment.[13] The gut consists of a central tube occupying the full length of the lobopodian's trunk,[14] which does not change much in width - at least not systematically. This may be surrounded by serially repeated[14] kidney-shaped diverticulae.[15] In some specimens, parts of the lobopodian gut can be preserved in three dimensions. This cannot result from phosphatisation, which is usually responsible for 3-D gut preservation,[16] for the phosphate content of the guts is under 1%; the contents comprise quartz and muscovite.[13] The gut of the representative Paucipodia is variable in width, being widest at the centre of the body. Its position in the body cavity is only loosely fixed, so flexibility is possible.
Diversity
During the Cambrian lobopodians displayed a substantial degree of biodiversity. One species is known from each of the Ordovician and Silurian periods,[5][17] with a few more known from the Carboniferous (Mazon Creek) — this represents the paucity of exceptional lagerstatten in post-Cambrian deposits.
Phylogeny
Budd sees the Lobopodia as representing a basal grade from which the phyla Onychophora and Arthropoda arose, with Aysheaia comparable to the ancestral plan, and with forms like Kerygmachaela and Pambdeleurion representing a transition that, via dinocaridids to arthropods, would lead to an arthropod body plan.[18] Aysheaia's surface ornamentation, if homologous with palaeoscolecid sclerites, may represent a deeper link connecting it with cycloneuralian-like outgroups.[18]
Budd sees the origin of armature in the armoured lobopodians as (originally) associated with muscle attachment points — as an adaptation to allow finer muscular control of the longer limbs seen in this group.[18] In Budd's view, armoured lobopodians represent a clade.[18]
More recent studies indicate that all three panarthropod phyla have lobopodians in their stem lineages. Lobopodians are thus paraphyletic, and include the common ancestor of Euarthropoda, Onychophora and Tardigrada. The most recent lobopodian phylogeny is displayed here.
References
- ↑ Haug, J. T.; Mayer, G.; Haug, C.; Briggs, D. E. G. (2012). "A Carboniferous Non-Onychophoran Lobopodian Reveals Long-Term Survival of a Cambrian Morphotype". Current Biology. 22: 1673–1675. doi:10.1016/j.cub.2012.06.066.
- ↑ Snodgrass, R.E. 1938. Evolution of the Annelida, Onychophora, and Arthropoda. Smithsonian Miscellaneous Collections, 97(6): 1-159
- ↑ BUDD, G. E. 1998. The morphology and phylogenetic significance of ~Kerygmachela kierkegaardi~ Budd (Buen Formation, Lower Cambrian, N Greenland). Transactions of the Royal Society of Edinburgh: Earth Sciences, 89, 249–290.
- ↑ Van Roy, P.; Orr, P. J.; Botting, J. P.; Muir, L. A.; Vinther, J.; Lefebvre, B.; Hariri, K. E.; Briggs, D. E. G. (2010). "Ordovician faunas of Burgess Shale type". Nature. 465 (7295): 215–8. Bibcode:2010Natur.465..215V. doi:10.1038/nature09038. PMID 20463737.
- 1 2 Von Bitter, P. H.; Purnell, M. A.; Tetreault, D. K.; Stott, C. A. (2007). "Eramosa Lagerstätte—Exceptionally preserved soft-bodied biotas with shallow-marine shelly and bioturbating organisms (Silurian, Ontario, Canada)". Geology. 35 (10): 879. doi:10.1130/G23894A.1.
- ↑ Haug, J. T.; Mayer, G.; Haug, C.; Briggs, D. E. G. (2012). "A Carboniferous Non-Onychophoran Lobopodian Reveals Long-Term Survival of a Cambrian Morphotype". Current Biology. 22: 1673–1675. doi:10.1016/j.cub.2012.06.066.
- ↑ Caron, J. -B.; Smith, M. R.; Harvey, T. H. P. (2013). "Beyond the Burgess Shale: Cambrian microfossils track the rise and fall of hallucigeniid lobopodians.". Proceedings of the Royal Society B: Biological Sciences. 280 (1767): 20131613. doi:10.1098/rspb.2013.1613. PMC 3735267
 . PMID 23902914.
. PMID 23902914. - ↑ Topper, T. P.; Skovsted, C. B.; Peel, J. S.; Harper, D. A. T. (2013). "Moulting in the lobopodian Onychodictyonfrom the lower Cambrian of Greenland". Lethaia: n/a. doi:10.1111/let.12026.
- ↑ Gould, S.J. (1989). Wonderful Life: The Burgess Shale and the Nature of History. W.W. Norton & Company.
- ↑ Further information and references: See Hallucigenia
- 1 2 Smith, Martin R.; Ortega-Hernández, Javier (2014). "Hallucigenia's onychophoran-like claws and the case for Tactopoda". Nature. 514 (7522): 363–366. Bibcode:2014Natur.514..363S. doi:10.1038/nature13576. PMID 25132546.
- ↑ MA, X., HOU, X. and BERGSTRÖM, J. 2009. Morphology of ~Luolishania longicruris~ (Lower Cambrian, Chengjiang Lagerstätte, SW China) and the phylogenetic relationships within lobopodians. Arthropod Structure & Development, 38, 271–291.
- 1 2 3 4 5 6 7 Hou, Xian-Guang; Ma, Xiao-Ya; Zhao, Jie; Bergström, Jan (2004). "The lobopodian Paucipodia inermis from the Lower Cambrian Chengjiang fauna, Yunnan, China". Lethaia. 37 (3): 235–244. doi:10.1080/00241160410006555.
- 1 2 Jianni Liu; Degan Shu; Jian Han; Zhifei Zhang & Xingliang Zhang (2006). "A large xenusiid lobopod with complex appendages from the Lower Cambrian Chengjiang Lagerstätte" (PDF). Acta Palaeontol. Pol. 51 (2): 215–222. Retrieved 9 February 2011.
- ↑ Liu, J.; Shu, D.; Han, J.; Zhang, Z.; Zhang, X. (2007). "Morpho-anatomy of the lobopod Magadictyon cf. Haikouensis from the Early Cambrian Chengjiang Lagerstätte, South China". Acta Zoologica. 88 (4): 279–288. doi:10.1111/j.1463-6395.2007.00281.x.
- ↑ Butterfield, N. J. (2002). "Leanchoilia guts and the interpretation of three-dimensional structures in Burgess Shale-type fossils". Paleobiology. 28: 155–171. doi:10.1666/0094-8373(2002)028<0155:LGATIO>2.0.CO;2. ISSN 0094-8373.
- ↑ Whittle, R. J.; Gabbott, S. E.; Aldridge, R. J.; Theron, J. (2009). "An Ordovician Lobopodian from the Soom Shale Lagerstätte, South Africa". Palaeontology. 52 (3): 561–567. doi:10.1111/j.1475-4983.2009.00860.x.
- 1 2 3 4 Budd, G. E. (2001). "Why are arthropods segmented?". Evolution and Development. 3 (5): 332–42. doi:10.1046/j.1525-142X.2001.01041.x. PMID 11710765.