Kainate receptor

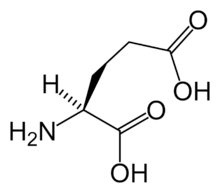
Kainate receptors, or KARs, are ionotropic receptors that respond to the neurotransmitter glutamate. They were first identified as a distinct receptor type through their selective activation by the agonist kainate, a drug first isolated from the red alga Digenea simplex. KARs are less understood than AMPA and NMDA receptors, the other ionotropic glutamate receptors. Postsynaptic kainate receptors are involved in excitatory neurotransmission. Presynaptic kainate receptors have been implicated in inhibitory neurotransmission by modulating release of the inhibitory neurotransmitter GABA through a presynaptic mechanism.
Structure
There are five types of kainate receptor subunits, GluR5 (GRIK1), GluR6 (GRIK2), GluR7 (GRIK3), KA1 (GRIK4) and KA2 (GRIK5), which are similar to AMPA and NMDA receptor subunits and can be arranged in different ways to form a tetramer, a four subunit receptor.[1] GluR5-7 can form homomers (ex. a receptor composed entirely of GluR5) and heteromers (ex. a receptor composed of both GluR5 and GluR6), however, KA1 and KA2 can only form functional receptors by combining with one of the GluR5-7 subunits. Since 2009 the kainate receptor subunits have been renamed to correspond with their gene name. Hence GluR5-7 are now GluK1-3 and KA1 and KA2 are GluK4 and GluK5 respectively.[2]
Each KAR subunit begins with a 400-residue extracellular N-terminal domain, which plays a key role in assembly, followed by the first segment of the neurotransmitter-binding cleft, called S1. This segment then passes through the cell membrane, forming the first of three membrane-spanning regions, M1. The M2 segment then begins on the cytoplasmic face of the membrane, pushes into the cell membrane about half way, and then dips back out to the cytoplasm. This segment, termed the "p loop," determines the calcium permeability of the receptor. M2 turns into M3, another transmembrane segment which emerges on the extracellular face to complete the neurotransmitter binding site (a portion called S2). M4 begins extracellularly, and passes again through the membrane into the cytoplasm, forming the C-terminal of the protein.
Conductance
The ion channel formed by kainate receptors is permeable to sodium and potassium ions. The single channel conductance of kainate receptor channels is similar to that of AMPA channels, at about 20 pS. However, rise and decay times for postsynaptic potentials generated by KARs are slower than for AMPA postsynaptic potentials. Their permeability to Ca2+ is usually very slight but varies with subunits and RNA editing at the tip of the p loop.[3]
Roles
Kainate receptors have both presynaptic and postsynaptic actions.[4] They have a somewhat more limited distribution in the brain than AMPA and NMDA receptors, and their function is less well defined. The convulsant kainic acid induces seizures, in part, by activation of kainate receptors containing the GluK2 subunit and also probably via AMPA receptors [5] Activation of kainate receptors containing the GluK1 subunit can also induce seizures but deletion of this subunit does not reduce seizure susceptibility to kainate or in other seizure models. Deletion of either GluK1 or GluK2 does not alter kindling epileptogenesis or the expression of kindled seizures.
Plasticity
Unlike AMPA receptors, kainate receptors play only a minor role in signaling at synapses.[6] Rather, kainate receptors may have a more subtle role in synaptic plasticity, affecting the likelihood that the postsynaptic cell will fire in response to future stimulation.[7][8] Activating kainate receptors in the presynaptic cell can affect the amount of neurotransmitters that are released[3][8][9] This effect may occur quickly and last for a long time,[9] and the effects of repetitive stimulation of KARs can be additive over time.[8]
Ligands
Agonists
- 5-Iodowillardiine
- ATPA
- Domoic acid
- Kainic acid
- LY-339,434
- SYM-2081
- Glutamate
Antagonists
- CNQX
- DNQX
- Ethanol
- NS102 - Selective of Kainate receptor over AMPA receptor
- Kynurenic acid - endogenous ligand
- Tezampanel
- UBP-302 - Highly selective for the kainate receptor
See also
References
- ↑ Dingledine R, Borges K, Bowie D, Traynelis SF (1999). "The glutamate receptor ion channels" (abstract). Pharmacol. Rev. 51 (1): 7–61. PMID 10049997.
- ↑ Collingridge GL, Olsen RW, Peters J, Spedding M (2009). "A nomenclature for ligand-gated ion channels". Neuropharmacology. 56 (1): 2–5. doi:10.1016/j.neuropharm.2008.06.063. PMC 2847504
 . PMID 18655795.
. PMID 18655795. - 1 2 Huettner JE (2003). "Kainate receptors and synaptic transmission". Prog. Neurobiol. 70 (5): 387–407. doi:10.1016/S0301-0082(03)00122-9. PMID 14511698.
- ↑ Contractor A, Mulle C, Swanson GT (2011). "Kainate receptors coming of age: milestones of two decades of research.". Trends Neurosci. 34 (3): 154–63. doi:10.1016/j.tins.2010.12.002. PMC 3051042. PMID 21256604.
- ↑ Fritsch B, Reis J, Gasior M, Kaminski RM, Rogawski MA (April 2014). "Role of GluK1 Kainate Receptors in Seizures, Epileptic Discharges, and Epileptogenesis". J. Neurosci. 34 (17): 5765–75. doi:10.1523/JNEUROSCI.5307-13.2014. PMID 24760837.
- ↑ Song I, Huganir RL (2002). "Regulation of AMPA receptors during synaptic plasticity". Trends Neurosci. 25 (11): 578–88. doi:10.1016/S0166-2236(02)02270-1. PMID 12392933.
- ↑ Contractor A, Swanson GT, Sailer A, O'Gorman S, Heinemann SF (2000). "Identification of the kainate receptor subunits underlying modulation of excitatory synaptic transmission in the CA3 region of the hippocampus" (abstract). J. Neurosci. 20 (22): 8269–78. PMID 11069933.
- 1 2 3 Mayer ML (2005). "Crystal structures of the GluR5 and GluR6 ligand binding cores: molecular mechanisms underlying kainate receptor selectivity". Neuron. 45 (4): 539–52. doi:10.1016/j.neuron.2005.01.031. PMID 15721240.
- 1 2 Schmitz D, Mellor J, Nicoll RA (2001). "Presynaptic kainate receptor mediation of frequency facilitation at hippocampal mossy fiber synapses". Science. 291 (5510): 1972–6. doi:10.1126/science.1057105. PMID 11239159.
External links
- Kainate Receptor at the US National Library of Medicine Medical Subject Headings (MeSH)