Fibroblast growth factor
| Fibroblast growth factor | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Crystal structure analysis of the FGF10-FGFR2b complex | |||||||||
| Identifiers | |||||||||
| Symbol | FGF | ||||||||
| Pfam | PF00167 | ||||||||
| Pfam clan | CL0066 | ||||||||
| InterPro | IPR002348 | ||||||||
| PROSITE | PDOC00220 | ||||||||
| SCOP | 1bas | ||||||||
| SUPERFAMILY | 1bas | ||||||||
| CDD | cd00058 | ||||||||
| |||||||||
Fibroblast growth factors, or FGFs, are a family of growth factors, with members involved in angiogenesis, wound healing, embryonic development and various endocrine signaling pathways. The FGFs are heparin-binding proteins and interactions with cell-surface-associated heparan sulfate proteoglycans have been shown to be essential for FGF signal transduction. FGFs are key players in the processes of proliferation and differentiation of wide variety of cells and tissues.
Families
In humans, 22 members of the FGF family have been identified, all of which are structurally related signaling molecules:[1][2][3]
- Members FGF1 through FGF10 all bind fibroblast growth factor receptors (FGFRs). FGF1 is also known as acidic, and FGF2 is also known as basic fibroblast growth factor.
- Members FGF11, FGF12, FGF13, and FGF14, also known as FGF homologous factors 1-4 (FHF1-FHF4), have been shown to have distinct functions compared to the FGFs. Although these factors possess remarkably similar sequence homology, they do not bind FGFRs and are involved in intracellular processes unrelated to the FGFs.[4] This group is also known as "iFGF".[5]
- Human FGF18 is involved in cell development and morphogenesis in various tissues including cartilage.[6]
- Human FGF20 was identified based on its homology to Xenopus FGF-20 (XFGF-20).[7][8]
- FGF15 through FGF23 were described later and functions are still being characterized. FGF15 is the mouse ortholog of human FGF19 (there is no human FGF15) and, where their functions are shared, they are often described as FGF15/19.[9] In contrast to the local activity of the other FGFs, FGF15/19, FGF21 and FGF23 have systemic effects.[9][10]
Receptors
The mammalian fibroblast growth factor receptor family has 4 members, FGFR1, FGFR2, FGFR3, and FGFR4. The FGFRs consist of three extracellular immunoglobulin-type domains (D1-D3), a single-span trans-membrane domain and an intracellular split tyrosine kinase domain. FGFs interact with the D2 and D3 domains, with the D3 interactions primarily responsible for ligand-binding specificity (see below). Heparan sulfate binding is mediated through the D3 domain. A short stretch of acidic amino acids located between the D1 and D2 domains has auto-inhibitory functions. This 'acid box' motif interacts with the heparan sulfate binding site to prevent receptor activation in the absence of FGFs.
Alternate mRNA splicing gives rise to 'b' and 'c' variants of FGFRs 1, 2 and 3. Through this mechanism seven different signaling FGFR sub-types can be expressed at the cell surface. Each FGFR binds to a specific subset of the FGFs. Similarly most FGFs can bind to several different FGFR subtypes. FGF1 is sometimes referred to as the 'universal ligand' as it is capable of activating all 7 different FGFRs. In contrast, FGF7 (keratinocyte growth factor, KGF) binds only to FGFR2b (KGFR).
The signaling complex at the cell surface is believed to be a ternary complex formed between two identical FGF ligands, two identical FGFR subunits, and either one or two heparan sulfate chains.
History
Fibroblast growth factor was found in pituitary extracts by Armelin in 1973[11] and then was also found in a cow brain extract by Gospodarowicz, et al., and tested in a bioassay that caused fibroblasts to proliferate (first published report in 1974).[12]
They then further fractionated the extract using acidic and basic pH and isolated two slightly different forms that were named "acidic fibroblast growth factor" (FGF1) and "basic fibroblast growth factor" (FGF2). These proteins had a high degree of amino acid identity but were determined to be distinct mitogens. Human FGF2 occurs in low molecular weight (LMW) and high molecular weight (HMW) isoforms.[13] LMW FGF2 is primarily cytoplasmic and functions in an autocrine manner, whereas HMW FGF2s are nuclear and exert activities through an intracrine mechanism.
Not long after FGF1 and FGF2 were isolated, another group isolated a pair of heparin-binding growth factors that they named HBGF-1 and HBGF-2, while a third group isolated a pair of growth factors that caused proliferation of cells in a bioassay containing blood vessel endothelium cells, which they called ECGF1 and ECGF2. These proteins were found to be identical to the acidic and basic FGFs described by Gospodarowicz, et al.
Functions
FGFs are multifunctional proteins with a wide variety of effects; they are most commonly mitogens but also have regulatory, morphological, and endocrine effects. They have been alternately referred to as "pluripotent" growth factors and as "promiscuous" growth factors due to their multiple actions on multiple cell types.[14][15] Promiscuous refers to the biochemistry and pharmacology concept of how a variety of molecules can bind to and elicit a response from single receptor. In the case of FGF, four receptor subtypes can be activated by more than twenty different FGF ligands. Thus the functions of FGFs in developmental processes include mesoderm induction, antero-posterior patterning,[7] limb development, neural induction and neural development,[16] and in mature tissues/systems angiogenesis, keratinocyte organization, and wound healing processes.
FGF is critical during normal development of both vertebrates and invertebrates and any irregularities in their function leads to a range of developmental defects.[17][18][19][20]
FGFs secreted by hypoblasts during avian gastrulation play a role in stimulating a Wnt signaling pathway that is involved in the differential movement of Koller's sickle cells during formation of the primitive streak.[21]
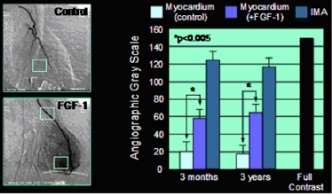
One important function of FGF1 and FGF2 is the promotion of endothelial cell proliferation and the physical organization of endothelial cells into tube-like structures. They thus promote angiogenesis, the growth of new blood vessels from the pre-existing vasculature. FGF1 and FGF2 are more potent angiogenic factors than vascular endothelial growth factor (VEGF) or platelet-derived growth factor (PDGF).[23] FGF1 has been shown in clinical experimental studies to induce angiogenesis in the heart.[22]
As well as stimulating blood vessel growth, FGFs are important players in wound healing. FGF1 and FGF2 stimulate angiogenesis and the proliferation of fibroblasts that give rise to granulation tissue, which fills up a wound space/cavity early in the wound-healing process. FGF7 and FGF10 (also known as Keratinocyte Growth Factors KGF and KGF2, respectively) stimulate the repair of injured skin and mucosal tissues by stimulating the proliferation, migration and differentiation of epithelial cells, and they have direct chemotactic effects on tissue remodeling.
During the development of the central nervous system, FGFs play important roles in neural stem cell proliferation, neurogenesis, axon growth, and differentiation. FGF signaling is important in promoting surface area growth of the developing cerebral cortex by reducing neuronal differentiation and hence permitting the self-renewal of cortical progenitor cells, known as radial glial cells,[24] and FGF2 has been used to induce artificial gyrification of the mouse brain.[25] Another FGF family member, FGF8, regulates the size and positioning of the functional areas of the cerebral cortex (Brodmann's Areas).[26][27] FGFs are also important for maintenance of the adult brain. Thus, FGFs are major determinants of neuronal survival both during development and during adulthood.[28] Adult neurogenesis within the hippocampus e.g. depends greatly on FGF2. In addition, FGF1 and FGF2 seem to be involved in the regulation of synaptic plasticity and processes attributed to learning and memory, at least in the hippocampus.
The 15 paracrine FGFs are secreted proteins that bind heparan sulfate and can, therefore, be bound to the extracellular matrix of tissues that contain heparan sulfate proteoglycans. This local action of FGF proteins is classified as paracrine signalling, most commonly through the JAK-STAT signaling pathway or the Receptor tyrosine kinase (RTK) pathway.
Members of the FGF19 subfamily (FGF15, FGF19, FGF21, and FGF23) bind less tightly to heparan sulfates, and so can act in an endocrine fashion on far-away tissues, such as intestine, liver, kidney, adipose, and bone.[9] For example:
- FGF15 and FGF19 (FGF15/19) are produced by intestinal cells but act on FGFR4-expressing liver cells to downregulate the key gene (CYP7A1) in the bile acid synthesis pathway.[29]
- FGF23 is produced by bone but acts on FGFR1-expressing kidney cells to regulate the synthesis of vitamin D and phosphate homeostasis.[30]
Structure
The crystal structures of FGF1 have been solved and found to be related to interleukin 1-beta. Both families have the same beta trefoil fold consisting of 12-stranded beta-sheet structure, with the beta-sheets are arranged in 3 similar lobes around a central axis, 6 strands forming an anti-parallel beta-barrel.[31][32][33] In general, the beta-sheets are well-preserved and the crystal structures superimpose in these areas. The intervening loops are less well-conserved - the loop between beta-strands 6 and 7 is slightly longer in interleukin-1 beta.
Clinical applications
Dysregulation of the FGF signalling system underlies a range of diseases, where inhibitors of FGF signalling have shown clinical efficacy.[34] Some FGF ligands (particularly FGF2) have been demonstrated to enhance tissue repair, e.g., skin burns and ulcers, in a range of clinical settings.[35]
See also
- Granulocyte-colony stimulating factor (G-CSF)
- Granulocyte-macrophage colony stimulating factor (GM-CSF)
- Nerve growth factor (NGF)
- Neurotrophins
- Erythropoietin (EPO)
- Thrombopoietin (TPO)
- Myostatin (GDF-8)
- Growth Differentiation factor-9 (GDF9)
- Gyrification
References
- ↑ Finklestein S.P.; Plomaritoglou A. (2001). "Growth factors". In Miller L.P., Hayes R.L. Co-edited by Newcomb J.K. Head Trauma: Basic, Preclinical, and Clinical Directions. New York: Wiley. pp. 165–187. ISBN 0-471-36015-5.
- ↑ Blaber M, DiSalvo J, Thomas KA (Feb 1996). "X-ray crystal structure of human acidic fibroblast growth factor". Biochemistry. 35 (7): 2086–94. doi:10.1021/bi9521755. PMID 8652550.
- ↑ Ornitz DM, Itoh N (2001). "Fibroblast growth factors". Genome Biology. 2 (3): reviews3005.1–reviews3005.12. doi:10.1186/gb-2001-2-3-reviews3005. PMC 138918
 . PMID 11276432.
. PMID 11276432. - ↑ Olsen SK, Garbi M, Zampieri N, Eliseenkova AV, Ornitz DM, Goldfarb M, Mohammadi M (Sep 2003). "Fibroblast growth factor (FGF) homologous factors share structural but not functional homology with FGFs". The Journal of Biological Chemistry. 278 (36): 34226–36. doi:10.1074/jbc.M303183200. PMID 12815063.
- ↑ Itoh N, Ornitz DM (Jan 2008). "Functional evolutionary history of the mouse Fgf gene family". Developmental Dynamics. 237 (1): 18–27. doi:10.1002/dvdy.21388. PMID 18058912.
- ↑ Moore EE, Bendele AM, Thompson DL, Littau A, Waggie KS, Reardon B, Ellsworth JL (Jul 2005). "Fibroblast growth factor-18 stimulates chondrogenesis and cartilage repair in a rat model of injury-induced osteoarthritis". Osteoarthritis and Cartilage / OARS, Osteoarthritis Research Society. 13 (7): 623–631. doi:10.1016/j.joca.2005.03.003. PMID 15896984.
- 1 2 Koga C, Adati N, Nakata K, Mikoshiba K, Furuhata Y, Sato S, Tei H, Sakaki Y, Kurokawa T, Shiokawa K, Yokoyama KK (Aug 1999). "Characterization of a novel member of the FGF family, XFGF-20, in Xenopus laevis". Biochemical and Biophysical Research Communications. 261 (3): 756–65. doi:10.1006/bbrc.1999.1039. PMID 10441498.
- ↑ Kirikoshi H, Sagara N, Saitoh T, Tanaka K, Sekihara H, Shiokawa K, Katoh M (Aug 2000). "Molecular cloning and characterization of human FGF-20 on chromosome 8p21.3-p22". Biochemical and Biophysical Research Communications. 274 (2): 337–43. doi:10.1006/bbrc.2000.3142. PMID 10913340.
- 1 2 3 Potthoff MJ, Kliewer SA, Mangelsdorf DJ (Feb 2012). "Endocrine fibroblast growth factors 15/19 and 21: from feast to famine". Genes & Development. 26 (4): 312–324. doi:10.1101/gad.184788.111. PMC 3289879. PMID 22302876.
- ↑ Fukumoto S (Mar 2008). "Actions and mode of actions of FGF19 subfamily members". Endocrine Journal. 55 (1): 23–31. doi:10.1507/endocrj.KR07E-002. PMID 17878606.
- ↑ Armelin HA (Sep 1973). "Pituitary extracts and steroid hormones in the control of 3T3 cell growth". Proceedings of the National Academy of Sciences of the United States of America. 70 (9): 2702–6. Bibcode:1973PNAS...70.2702A. doi:10.1073/pnas.70.9.2702. PMC 427087. PMID 4354860.
- ↑ Gospodarowicz D (May 1974). "Localisation of a fibroblast growth factor and its effect alone and with hydrocortisone on 3T3 cell growth". Nature. 249 (453): 123–7. Bibcode:1974Natur.249..123G. doi:10.1038/249123a0. PMID 4364816.
- ↑ Arese M, Chen Y, Florkiewicz RZ, Gualandris A, Shen B, Rifkin DB (May 1999). "Nuclear activities of basic fibroblast growth factor: potentiation of low-serum growth mediated by natural or chimeric nuclear localization signals". Molecular Biology of the Cell. 10 (5): 1429–44. doi:10.1091/mbc.10.5.1429. PMC 25296. PMID 10233154.
- ↑ Vlodavsky I, Korner G, Ishai-Michaeli R, Bashkin P, Bar-Shavit R, Fuks Z (Nov 1990). "Extracellular matrix-resident growth factors and enzymes: possible involvement in tumor metastasis and angiogenesis". Cancer Metastasis Reviews. 9 (3): 203–26. doi:10.1007/BF00046361. PMID 1705486.
- ↑ Green PJ, Walsh FS, Doherty P (Aug 1996). "Promiscuity of fibroblast growth factor receptors". BioEssays. 18 (8): 639–46. doi:10.1002/bies.950180807. PMID 8760337.
- ↑ Böttcher RT, Niehrs C (Feb 2005). "Fibroblast growth factor signaling during early vertebrate development". Endocrine Reviews. 26 (1): 63–77. doi:10.1210/er.2003-0040. PMID 15689573.
- ↑ Amaya E, Musci TJ, Kirschner MW (Jul 1991). "Expression of a dominant negative mutant of the FGF receptor disrupts mesoderm formation in Xenopus embryos". Cell. 66 (2): 257–270. doi:10.1016/0092-8674(91)90616-7. PMID 1649700.
- ↑ Borland CZ, Schutzman JL, Stern MJ (Dec 2001). "Fibroblast growth factor signaling in Caenorhabditis elegans". BioEssays. 23 (12): 1120–1130. doi:10.1002/bies.10007. PMID 11746231.
- ↑ Coumoul X, Deng CX (Nov 2003). "Roles of FGF receptors in mammalian development and congenital diseases". Birth Defects Research. Part C, Embryo Today. 69 (4): 286–304. doi:10.1002/bdrc.10025. PMID 14745970.
- ↑ Sutherland D, Samakovlis C, Krasnow MA (Dec 1996). "branchless encodes a Drosophila FGF homolog that controls tracheal cell migration and the pattern of branching". Cell. 87 (6): 1091–1101. doi:10.1016/S0092-8674(00)81803-6. PMID 8978613.
- ↑ Gilbert SF. Developmental Biology. 10th edition. Sunderland (MA): Sinauer Associates; 2014. Early Development in Birds. Print
- 1 2 Stegmann, TJ (May 1999). "New approaches to coronary heart disease: induction of neovascularisation by growth factors". BioDrugs. 11 (5): 301–8. doi:10.2165/00063030-199911050-00002. PMID 18031140.
- ↑ Cao R, Bråkenhielm E, Pawliuk R, Wariaro D, Post MJ, Wahlberg E, Leboulch P, Cao Y (May 2003). "Angiogenic synergism, vascular stability and improvement of hind-limb ischemia by a combination of PDGF-BB and FGF-2". Nature Medicine. 9 (5): 604–13. doi:10.1038/nm848. PMID 12669032.
- ↑ Rash BG, Lim HD, Breunig JJ, Vaccarino FM (Oct 2011). "FGF signaling expands embryonic cortical surface area by regulating Notch-dependent neurogenesis". The Journal of Neuroscience. 31 (43): 15604–17. doi:10.1523/JNEUROSCI.4439-11.2011. PMID 22031906.
- ↑ Rash BG, Tomasi S, Lim HD, Suh CY, Vaccarino FM (Jun 2013). "Cortical gyrification induced by fibroblast growth factor 2 in the mouse brain". The Journal of Neuroscience. 33 (26): 10802–14. doi:10.1523/JNEUROSCI.3621-12.2013. PMC 3693057. PMID 23804101.
- ↑ Fukuchi-Shimogori T, Grove EA (Nov 2001). "Neocortex patterning by the secreted signaling molecule FGF8". Science. 294 (5544): 1071–4. Bibcode:2001Sci...294.1071F. doi:10.1126/science.1064252. PMID 11567107.
- ↑ Garel S, Huffman KJ, Rubenstein JL (May 2003). "Molecular regionalization of the neocortex is disrupted in Fgf8 hypomorphic mutants". Development. 130 (9): 1903–14. doi:10.1242/dev.00416. PMID 12642494.
- ↑ Reuss B, von Bohlen und Halbach O (Aug 2003). "Fibroblast growth factors and their receptors in the central nervous system". Cell and Tissue Research. 313 (2): 139–157. doi:10.1007/s00441-003-0756-7. PMID 12845521.
- ↑ Jones SA (2012). "Physiology of FGF15/19". Advances in Experimental Medicine and Biology. Advances in Experimental Medicine and Biology. 728: 171–82. doi:10.1007/978-1-4614-0887-1_11. ISBN 978-1-4614-0886-4. PMID 22396169.
- ↑ Razzaque MS (Nov 2009). "The FGF23-Klotho axis: endocrine regulation of phosphate homeostasis". Nature Reviews. Endocrinology. 5 (11): 611–9. doi:10.1038/nrendo.2009.196. PMC 3107967. PMID 19844248.
- ↑ Murzin AG, Lesk AM, Chothia C (Jan 1992). "beta-Trefoil fold. Patterns of structure and sequence in the Kunitz inhibitors interleukins-1 beta and 1 alpha and fibroblast growth factors". Journal of Molecular Biology. 223 (2): 531–43. doi:10.1016/0022-2836(92)90668-A. PMID 1738162.
- ↑ Eriksson AE, Cousens LS, Weaver LH, Matthews BW (Apr 1991). "Three-dimensional structure of human basic fibroblast growth factor". Proceedings of the National Academy of Sciences of the United States of America. 88 (8): 3441–5. Bibcode:1991PNAS...88.3441E. doi:10.1073/pnas.88.8.3441. PMC 51463. PMID 1707542.
- ↑ Gimenez-Gallego G, Rodkey J, Bennett C, Rios-Candelore M, DiSalvo J, Thomas K (Dec 1985). "Brain-derived acidic fibroblast growth factor: complete amino acid sequence and homologies". Science. 230 (4732): 1385–8. Bibcode:1985Sci...230.1385G. doi:10.1126/science.4071057. PMID 4071057.
- ↑ Carter EP, Fearon AE, Grose RP (Apr 2015). "Careless talk costs lives: fibroblast growth factor receptor signalling and the consequences of pathway malfunction". Trends Cell Biol. 25 (4): 221–33. doi:10.1016/j.tcb.2014.11.003. PMID 25467007.
- ↑ Nunes QM, Li Y, Sun C, Kinnunen TK, Fernig DG (Jan 2016). "Fibroblast growth factors as tissue repair and regeneration therapeutics". PeerJ. 4: e1535. doi:10.7717/peerj.1535. PMC 4715458. PMID 26793421.
External links
| Wikimedia Commons has media related to Fibroblast growth factor. |
- Fibroblast Growth Factors at the US National Library of Medicine Medical Subject Headings (MeSH)
- FGF1 in Cosmetic Products
This article incorporates text from the public domain Pfam and InterPro IPR002348