Feathered dinosaur

A feathered dinosaur is any species of dinosaur possessing feathers. For over 150 years, since scientific research began on dinosaurs in the early 1800s, dinosaurs were generally believed to be related to the reptile family; the word "dinosaur", coined in 1842 by paleontologist Richard Owen, comes from the Greek for "terrible lizard". This view began to shift during the so-called dinosaur renaissance in scientific research in the late 1960s, and by the mid-1990s significant evidence had emerged that dinosaurs are much more closely related to birds. In fact, birds are now believed to have descended directly from the theropod group of dinosaurs,[1] and are thus classified as dinosaurs themselves, meaning that any modern bird can be considered a feathered dinosaur, since all modern birds possess feathers (with the exception of a few artificially selected chickens).[2]
Among extinct dinosaurs, feathers or feather-like integument have been discovered on dozens of genera via both direct and indirect fossil evidence. The vast majority of feather discoveries have been for coelurosaurian theropods. However, integument has also been discovered on at least three ornithischians, raising the likelihood that proto-feathers were also present in earlier dinosaurs, and perhaps even a more ancestral animal, in light of the pycnofibers of pterosaurs. Crocodilians also possess beta keratin very similar to those of birds, which suggests that they evolved from a common ancestral gene.[3]
History of research
Early
.jpg)
Shortly after the 1859 publication of Charles Darwin's On the Origin of Species, British biologist Thomas Henry Huxley proposed that birds were descendants of dinosaurs. He compared the skeletal structure of Compsognathus, a small theropod dinosaur, and the 'first bird' Archaeopteryx lithographica (both of which were found in the Upper Jurassic Bavarian limestone of Solnhofen). He showed that, apart from its hands and feathers, Archaeopteryx was quite similar to Compsognathus. Thus Archaeopteryx represents a transitional fossil. In 1868 he published On the Animals which are most nearly intermediate between Birds and Reptiles, making the case.[4][5] The first restoration of a feathered dinosaur was Thomas Henry Huxley's depiction in 1876 of a feathered Compsognathus to accompany a lecture on the evolution of birds he delivered in New York in which he speculated that the aforementioned dinosaur might have been in possession of feathers. [6] The leading dinosaur expert of the time, Richard Owen, disagreed, claiming Archaeopteryx as the first bird outside dinosaur lineage. For the next century, claims that birds were dinosaur descendants faded, with more popular bird-ancestry hypotheses including 'crocodylomorph' and 'thecodont' ancestors, rather than dinosaurs or other archosaurs.
In 1969, John Ostrom described Deinonychus antirrhopus, a theropod that he had discovered in Montana in 1964 and whose skeletal resemblance to birds seemed unmistakable. Ostrom became a leading proponent of the theory that birds are direct descendants of dinosaurs. Further comparisons of bird and dinosaur skeletons, as well as cladistic analysis strengthened the case for the link, particularly for a branch of theropods called maniraptors. Skeletal similarities include the neck, the pubis, the wrists (semi-lunate carpal), the 'arms' and pectoral girdle, the shoulder blade, the clavicle and the breast bone. In all, over a hundred distinct anatomical features are shared by birds and theropod dinosaurs.
Other researchers drew on these shared features and other aspects of dinosaur biology and began to suggest that at least some theropod dinosaurs were feathered. Gregory S. Paul was probably among the first paleoartists to depict non-avian maniraptoran dinosaurs with feathers and protofeathers, starting in the late 1970s.
By the 1990s, most paleontologists considered birds to be surviving dinosaurs and referred to 'non-avian dinosaurs' (all extinct), to distinguish them from birds (Avialae). Before the discovery of feathered dinosaurs, the evidence was limited to Huxley and Ostrom's comparative anatomy. Some mainstream ornithologists, including Smithsonian Institution curator Storrs L. Olson, disputed the links, specifically citing the lack of fossil evidence for feathered dinosaurs.
Fossil discoveries

After a century of hypotheses without conclusive evidence, well-preserved fossils of feathered dinosaurs were discovered during the 1990s, and more continue to be found. The fossils were preserved in a Lagerstätte — a sedimentary deposit exhibiting remarkable richness and completeness in its fossils — in Liaoning, China. The area had repeatedly been smothered in volcanic ash produced by eruptions in Inner Mongolia 124 million years ago, during the Early Cretaceous epoch. The fine-grained ash preserved the living organisms that it buried in fine detail. The area was teeming with life, with millions of leaves, angiosperms (the oldest known), insects, fish, frogs, salamanders, mammals, turtles, and lizards discovered to date.
The most important discoveries at Liaoning have been a host of feathered dinosaur fossils, with a steady stream of new finds filling in the picture of the dinosaur–bird connection and adding more to theories of the evolutionary development of feathers and flight. Turner et al. (2007) reported quill knobs from an ulna of Velociraptor mongoliensis, and these are strongly correlated with large and well-developed secondary feathers.[7]

Behavioural evidence, in the form of an oviraptorosaur on its nest, showed another link with birds. Its forearms were folded, like those of a bird.[8] Although no feathers were preserved, it is likely that these would have been present to insulate eggs and juveniles.[9]
Not all of the Chinese fossil discoveries proved valid however. In 1999, a supposed fossil of an apparently feathered dinosaur named Archaeoraptor liaoningensis, found in Liaoning Province, northeastern China, turned out to be a forgery. Comparing the photograph of the specimen with another find, Chinese paleontologist Xu Xing came to the conclusion that it was composed of two portions of different fossil animals. His claim made National Geographic review their research and they too came to the same conclusion.[10] The bottom portion of the "Archaeoraptor" composite came from a legitimate feathered dromaeosaurid now known as Microraptor, and the upper portion from a previously-known primitive bird called Yanornis.
In 2011, samples of amber were discovered to contain preserved feathers from 75 to 80 million years ago during the Cretaceous era, with evidence that they were from both dinosaurs and birds. Initial analysis suggests that some of the feathers were used for insulation, and not flight.[11][12] More complex feathers were revealed to have variations in coloration similar to modern birds, while simpler protofeathers were predominantly dark. Only 11 specimens are currently known. The specimens are too rare to be broken open to study their melanosomes, but there are plans for using non-destructive high-resolution X-ray imaging.[13]
Current knowledge
List of non-avian dinosaur species preserved with evidence of feathers


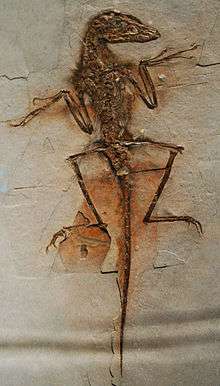

A number of non-avian[14] dinosaurs are now known to have been feathered. Direct evidence of feathers exists for the following species, listed in the order currently accepted evidence was first published. In all examples, the evidence described consists of feather impressions, except those genera inferred to have had feathers based on skeletal or chemical evidence, such as the presence of quill knobs (the anchor points for wing feathers on the forelimb) or a pygostyle (the fused vertebrae at the tail tip which often supports large feathers).
- Avimimus portentosus (inferred 1987: ulnar ridge)[15][16]
- Sinosauropteryx prima (1996)[17]
- Protarchaeopteryx robusta (1997)[18]
- GMV 2124 (1997)[19]
- Caudipteryx zoui (1998)[20]
- Rahonavis ostromi (inferred 1998: quill knobs; possibly avialan[21])[22]
- Shuvuuia deserti (1999)[23]
- Beipiaosaurus inexpectus (1999)[24]
- Sinornithosaurus millenii (1999)[25]
- Caudipteryx dongi (2000)[26]
- Caudipteryx sp. (2000)[27]
- Microraptor zhaoianus (2000)[28]
- Nomingia gobiensis (inferred 2000: pygostyle)[29]
- Psittacosaurus sp.? (2002)[30]
- Scansoriopteryx heilmanni (2002; possibly avialan)[31]
- Yixianosaurus longimanus (2003)[32]
- Dilong paradoxus (2004)[33]
- Pedopenna daohugouensis (2005; possibly avialan[34])[35]
- Jinfengopteryx elegans (2005)[36][37]
- Juravenator starki (2006)[38][39]
- Sinocalliopteryx gigas (2007)[40]
- Velociraptor mongoliensis (inferred 2007: quill knobs)[7]
- Epidexipteryx hui (2008; possibly avialan)[41]
- Similicaudipteryx yixianensis (inferred 2008: pygostyle; confirmed 2010)[42][43]
- Anchiornis huxleyi (2009; possibly avialan)[44]
- Tianyulong confuciusi? (2009)[45]
- Xiaotingia zhengi (2011; possibly avialan)[46]
- Yutyrannus huali (2012)[47]
- Sciurumimus albersdoerferi (2012)[48]
- Ornithomimus edmontonicus (2012)[49]
- Ningyuansaurus wangi (2012)[50]
- Eosinopteryx brevipenna (2013; possibly avialan)[51]
- Jianchangosaurus yixianensis (2013)[52]
- Aurornis xui (2013; possibly avialan)[53]
- Changyuraptor yangi (2014)[54]
- Kulindadromeus zabaikalicus? (2014)[55]
- Citipati osmolskae (inferred 2014: pygostyle)[56]
- Conchoraptor gracilis (inferred 2014: pygostyle)[56]
- Deinocheirus mirificus (inferred 2014: pygostyle)[57]
- Yi qi (2015)[58]
- Zhenyuanlong suni (2015)[59]
- Dakotaraptor steini (inferred 2015: quill knobs)[60]
- Apatoraptor pennatus (inferred 2016: quill knobs)[61]
- Note that the filamentous structures in some ornithischian dinosaurs (Psittacosaurus, Tianyulong and Kulindadromeus) and the pycnofibres found in some pterosaurs may or may not be homologous with the feathers of theropods.[45][62]
Primitive feather types
Integumentary structures that gave rise to the feathers of birds are seen in the dorsal spines of reptiles and fish. A similar stage in their evolution to the complex coats of birds and mammals can be observed in living reptiles such as iguanas and Gonocephalus agamids. Feather structures are thought to have proceeded from simple hollow filaments through several stages of increasing complexity, ending with the large, deeply rooted feathers with strong pens (rachis), barbs and barbules that birds display today.[63]
According to Prum's (1999) proposed model, at stage I, the follicle originates with a cylindrical epidermal depression around the base of the feather papilla. The first feather resulted when undifferentiated tubular follicle collar developed out of the old keratinocytes being pushed out. At stage II, the inner, basilar layer of the follicle collar differentiated into longitudinal barb ridges with unbranched keratin filaments, while the thin peripheral layer of the collar became the deciduous sheath, forming a tuft of unbranched barbs with a basal calamus. Stage III consists of two developmental novelties, IIIa and IIIb, as either could have occurred first. Stage IIIa involves helical displacement of barb ridges arising within the collar. The barb ridges on the anterior midline of the follicle fuse together, forming the rachis. The creation of a posterior barb locus follows, giving an indeterminate number of barbs. This resulted in a feather with a symmetrical, primarily branched structure with a rachis and unbranched barbs. In stage IIIb, barbules paired within the peripheral barbule plates of the barb ridges, create branched barbs with rami and barbules. This resulting feather is one with a tuft of branched barbs without a rachis. At stage IV, differentiated distal and proximal barbules produce a closed, pennaceous vane. A closed vane develops when pennulae on the distal barbules form a hooked shape to attach to the simpler proximal barbules of the adjacent barb. Stage V developmental novelties gave rise to additional structural diversity in the closed pennaceous feather. Here, asymmetrical flight feathers, bipinnate plumulaceous feathers, filoplumes, powder down, and bristles evolved.[64]
Some evidence suggests that the original function of simple feathers was insulation. In particular, preserved patches of skin in large, derived, tyrannosauroids show scutes, while those in smaller, more primitive, forms show feathers. This may indicate that the larger forms had complex skins, with both scutes and filaments, or that tyrannosauroids may be like rhinos and elephants, having filaments at birth and then losing them as they developed to maturity.[33] An adult Tyrannosaurus rex weighed about as much as an African elephant. If large tyrannosauroids were endotherms, they would have needed to radiate heat efficiently, and feathers would have interfered with this.[65]
There is an increasing body of evidence that supports the display hypothesis, which states that early feathers were colored and increased reproductive success.[66][67] Coloration could have provided the original adaptation of feathers, implying that all later functions of feathers, such as thermoregulation and flight, were co-opted.[66] This hypothesis has been supported by the discovery of pigmented feathers in multiple species.[68][69][70] Supporting the display hypothesis is the fact that fossil feathers have been observed in a ground-dwelling herbivorous dinosaur clade, making it unlikely that feathers functioned as predatory tools or as a means of flight.[71] Additionally, some specimens have iridescent feathers.[72] Pigmented and iridescent feathers may have provided greater attractiveness to mates, providing enhanced reproductive success when compared to non-colored feathers. Current research shows that it is plausible that theropods would have had the visual acuity necessary to see the displays. In a study by Stevens (2006), the binocular field of view for Velociraptor has been estimated to be 55 to 60 degrees, which is about that of modern owls. Visual acuity for Tyrannosaurus has been predicted to be anywhere from about that of humans to 13 times that of humans.[73] However, as both Velociraptor and Tyrannosaurus have a rather extended evolutionary relationship with the more basal theropods, it is unclear how much of this visual acuity data can be extrapolated.
The fact that precursors of feathers appeared and then were co-opted for insulation is already present in Gould and Vrba, 1982.[74] The reason why such precursors appeared could be explained by a theory which is based on metabolic issue. Feathers are made of protein and contain substantial amounts of certain amino acids, especially cysteine. The protein complex at the base of the composition of the feather is keratin, which has disulfide bonds between amino acids that confer unique properties of stability and elasticity. The metabolism of amino acids containing sulfur proved to be toxic to the organism. If the sulfur amino acids are not catabolized at the final products of urea or uric acid but used for the synthesis of keratin instead, the release of hydrogen sulfide is extremely reduced or avoided. For an organism whose metabolism works at high internal temperatures of 40 °C or greater can be extremely important to prevent the excess production of hydrogen sulfide. This hypothesis could be consistent with the need for high metabolic rate of theropod dinosaurs.[75][76]
It is not known with certainty at what point in archosaur phylogeny the earliest simple "protofeathers" arose, or whether they arose once or independently multiple times. Filamentous structures are clearly present in pterosaurs, and long, hollow quills have been reported in specimens of the ornithischian dinosaurs Psittacosaurus and Tianyulong.[30][45] In 2009, Xu et al. noted that the hollow, unbranched, stiff integumentary structures found on a specimen of Beipiaosaurus were strikingly similar to the integumentary structures of Psittacosaurus and pterosaurs. They suggested that all of these structures may have been inherited from a common ancestor much earlier in the evolution of archosaurs, possibly in an ornithodire from the Middle Triassic or earlier.[77] More recently, findings in Russia of the basal neornithischian Kulindadromeus report that although the lower leg and tail seemed to be scaled, "varied integumentary structures were found directly associated with skeletal elements, supporting the hypothesis that simple filamentous feathers, as well as compound feather-like structures comparable to those in theropods, were widespread amongst the whole dinosaur clade."[78]
Display feathers are also known from dinosaurs that are very primitive members of the bird lineage, or Avialae. The most primitive example is Epidexipteryx, which had a short tail with extremely long, ribbon-like feathers. Oddly enough, the fossil does not preserve wing feathers, suggesting that Epidexipteryx was either secondarily flightless, or that display feathers evolved before flight feathers in the bird lineage.[41] Plumaceous feathers are found in nearly all lineages of Theropoda common in the northern hemisphere, and pennaceous feathers are attested as far down the tree as the Ornithomimosauria. The fact that only adult Ornithomimus had wing-like structures suggests that pennaceous feathers evolved for mating displays.[79]
Phylogeny and the inference of feathers in other dinosaurs
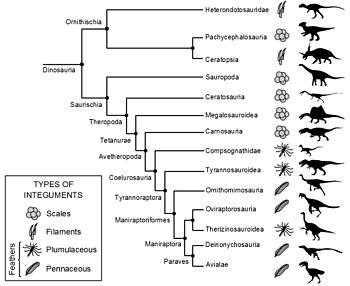
Fossil feather impressions are extremely rare and they require exceptional preservation conditions to form. Therefore, only a few feathered dinosaur genera have been identified. All fossil feather specimens have been found to show certain similarities. Due to these similarities and through developmental research, almost all scientists agree that feathers could only have evolved once in dinosaurs. Feathers would then have been passed down to all later, more derived species, unless some lineages lost feathers secondarily. If a dinosaur falls at a point on an evolutionary tree within the known feather-bearing lineages, then its ancestors had feathers, and it is quite possible that it did as well. This technique, called phylogenetic bracketing, can also be used to infer the type of feathers a species may have had, since the developmental history of feathers is now reasonably well-known. All feathered species had filamentaceous or plumaceous (downy) feathers, with pennaceous feathers found among the more bird-like groups. The following cladogram is adapted from Godefroit et al., 2013.[80]
Grey denotes a clade that is not known to contain any feathered specimen at the time of writing (although this does not imply that members of the clade lacked feathers).
| Neotheropoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
See also
References
- ↑ Brown, J.W.; Van Tuinen, M. (2011). "Evolving Perceptions on the Antiquity of the Modern Avian Tree, in Living Dinosaurs". The Evolutionary History of Modern Birds. John Wiley & Sons LtD: 306–324. doi:10.1002/9781119990475.ch12. ISBN 9781119990475.
- ↑ "Bald chicken 'needs no plucking'". BBC. Israel. May 21, 2002. Retrieved March 16, 2015.
- ↑ Greenwold, M,; Sawyer, R, (2013), "Molecular evolution and expression of archosaurian β-keratins: Diversification and expansion of archosaurian β-keratins and the origin of feather β-keratins", Journal of Experimental Zoology, 320 (6): 393–405, doi:10.1002/jez.b.22514, PMID 23744807, 22514
- ↑ Huxley, T.H. (1868). "On the animals which are most nearly intermediate between birds and reptiles". Annals and Magazine of Natural History. 4th. 2: 66–75.
- ↑ Foster, Michael; Lankester, E. Ray 1898–1903. The scientific memoirs of Thomas Henry Huxley. 4 vols and supplement. London: Macmillan.
- ↑ Huxley, Thomas Henry (1877). "The Hypothesis of Evolution. The Neutral and the Favourable Evidence.". In Huxley, Thomas Henry (ed.). American Addresses, With A Lecture On Biology. New York: D. Appleton And Company.
- 1 2 Turner, A.H.; Makovicky, P.J.; Norell, M.A. (2007). "Feather quill knobs in the dinosaur Velociraptor" (pdf). Science. 317 (5845): 1721. Bibcode:2007Sci...317.1721T. doi:10.1126/science.1145076. PMID 17885130.
- ↑ Norell M.A., Clark J.M., Chiappe L.M., Dashzeveg D.; Clark; Chiappe; Dashzeveg (1995). "A nesting dinosaur". Nature. 378 (6559): 774–776. Bibcode:1995Natur.378..774N. doi:10.1038/378774a0.
- ↑ Hopp, Thomas J., Orsen, Mark J. (2004) "Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Chapter 11. Dinosaur Brooding Behavior and the Origin of Flight Feathers" Currie, Koppelhaus, Shugar, Wright. Indiana University Press. Bloomington, IN. USA.
- ↑ "Transcript: The Dinosaur that Fooled the World". BBC. Retrieved 22 December 2006.
- ↑ Bock, Walter J. "American Zoologist". American Zoologist. 40: 478–485. doi:10.1668/0003-1569(2000)040[0478:ehotoo]2.0.co;2.
- ↑ Emily Chung (12 September 2011). "Dinosaur feathers found in Alberta amber". CBC. Retrieved 16 September 2011.
- ↑ Brian Switek (15 September 2011). "Amber inclusions showcase prehistoric feathers". Nature News. doi:10.1038/news.2011.539. Retrieved 22 September 2011.
- ↑ Stephen L. Brusatte, Graeme T. Lloyd, Steve C. Wang, Mark A. Norell (2014). "Gradual assembly of avian body plan culminated in rapid rates of evolution across the dinosaur-bird transition". Current Biology. 24 (20): 2386–2392. doi:10.1016/j.cub.2014.08.034.
- ↑ Kurzanov, S.M. (1987). "Avimimidae and the problem of the origin of birds." Transactions of the Joint Soviet-Mongolian Paleontological Expedition, 31: 5-92. [in Russian]
- ↑ Chiappe, L.M. and Witmer, L.M. (2002). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press, 536 pp. ISBN 0-520-20094-2
- ↑ Ji Q., Ji S. (1996). "On discovery of the earliest bird fossil in China and the origin of birds". Chinese Geology. 10 (233): 30–33.
- ↑ Ji, Q., and Ji, S. (1997). "A Chinese archaeopterygian, Protarchaeopteryx gen. nov." Geological Science and Technology (Di Zhi Ke Ji), 238: 38-41. Translated By Will Downs Bilby Research Center Northern Arizona University January 2001
- ↑ Ji, Q.; Ji, S. (1997). "Advances in Sinosauropteryx research". Chinese Geology. 7: 30–32.
- ↑ Currie, Philip J.; Qiang, Ji; Norell, Mark A.; Shu-An, Ji (1998). "Two feathered dinosaurs from northeastern China". Nature. 393 (6687): 753–761. Bibcode:1998Natur.393..753Q. doi:10.1038/31635.
- ↑ Agnolin, F.L.; Novas, F.E. (2011). "Unenlagiid theropods: are they members of the Dromaeosauridae (Theropoda, Maniraptora)?" (PDF). Anais da Academia Brasileira de Ciências. 83: 117–162. doi:10.1590/S0001-37652011000100008. Retrieved 23 April 2011.
- ↑ Forster, Catherine A.; Sampson, Scott D.; Chiappe, Luis M.; Krause, David W. (1998a). "The Theropod Ancestry of Birds: New Evidence from the Late Cretaceous of Madagascar". Science. 279 (5358): 1915–1919. Bibcode:1998Sci...279.1915F. doi:10.1126/science.279.5358.1915. PMID 9506938. (HTML abstract)
- ↑ Schweitzer, M.H.; Watt, J.A.; Avci, R.; Knapp, L.; Chiappe, L.; Norell, M.; Marshall, M. (1999). "Beta-keratin specific immunological reactivity in feather-like structures of the Cretaceous Alvarezsaurid,Shuvuuia deserti". Journal of Experimental Zoology. 285 (2): 146–57. doi:10.1002/(SICI)1097-010X(19990815)285:2<146::AID-JEZ7>3.0.CO;2-A. PMID 10440726.
- ↑ Xu, Xing; Tang, Zhi-lu; Wang, Xiao-lin (1999). "A therizinosauroid dinosaur with integumentary structures from China". Nature. 399 (6734): 350–354. Bibcode:1999Natur.399..350X. doi:10.1038/20670.
- ↑ Wu, Xiao-Chun; Xu, Xing; Wang, Xiao-Lin (1999). "A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China". Nature. 401 (6750): 262–266. Bibcode:1999Natur.401..262X. doi:10.1038/45769.
- ↑ Zhou, Z.; Wang, X. (2000). "A new species of Caudipteryx from the Yixian Formation of Liaoning, northeast China" (PDF). Vertebrata PalAsiatica. 38 (2): 113–130.
- ↑ Zhou, Z.; Wang, X.; Zhang, F.; Xu, X. (2000). "Important features of Caudipteryx - Evidence from two nearly complete new specimens" (PDF). Vertebrata PalAsiatica. 38 (4): 241–254.
- ↑ Xu, Xing; Zhou, Zhonghe; Wang, Xiaolin (2000). "The smallest known non-avian theropod dinosaur" (PDF). Nature. 408 (6813): 705–708. doi:10.1038/35047056. PMID 11130069.
- ↑ Barsbold R., Osmólska H., Watabe M., Currie P.J., Tsogtbaatar K. (2000). "New Oviraptorosaur (Dinosauria, Theropoda) From Mongolia: The First Dinosaur With A Pygostyle" (PDF). Acta Palaeontologica Polonica. 45 (2): 97–106.
- 1 2 Mayr, Gerald; Peters, Stefan; Plodowski, Gerhard; Vogel, Olaf (2002). "Bristle-like integumentary structures at the tail of the horned dinosaur Psittacosaurus". Naturwissenschaften. 89 (8): 361–365. Bibcode:2002NW.....89..361M. doi:10.1007/s00114-002-0339-6. PMID 12435037.
- ↑ Czerkas, S.A., and Yuan, C. (2002). "An arboreal maniraptoran from northeast China." Pp. 63-95 in Czerkas, S.J. (Ed.), Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum Journal 1. The Dinosaur Museum, Blanding, U.S.A. PDF abridged version
- ↑ Xu X.; Wang X.-L. (2003). "A new maniraptoran from the Early Cretaceous Yixian Formation of western Liaoning". Vertebrata PalAsiatica. 41 (3): 195–202.
- 1 2 Xu, X., Norell, M. A., Kuang, X., Wang, X., Zhao, Q., Jia, C. (2004). "Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids". Nature. 431 (7009): 680–684. Bibcode:2004Natur.431..680X. doi:10.1038/nature02855. PMID 15470426.
- ↑ Hone D.W.E., Tischlinger H., Xu X., Zhang F. (2010). Farke, Andrew Allen, ed. "The extent of the preserved feathers on the four-winged dinosaur Microraptor gui under ultraviolet light". PLoS ONE. 5 (2): e9223. Bibcode:2010PLoSO...5.9223H. doi:10.1371/journal.pone.0009223. PMC 2821398
 . PMID 20169153.
. PMID 20169153. - ↑ Xu, Xing; Zhang, Fucheng (2005). "A new maniraptoran dinosaur from China with long feathers on the metatarsus". Naturwissenschaften. 92 (4): 173–177. Bibcode:2005NW.....92..173X. doi:10.1007/s00114-004-0604-y. PMID 15685441.
- ↑ Ji Q., Ji S., Lu J., You H., Chen W., Liu Y., Liu Y. (2005). "First avialan bird from China (Jinfengopteryx elegans gen. et sp. nov.)". Geological Bulletin of China. 24 (3): 197–205.
- ↑ Turner, Alan H.; Pol, Diego; Clarke, Julia A.; Erickson, Gregory M.; Norell, Mark A. (2007). "A basal dromaeosaurid and size evolution preceding avian flight" (pdf). Science. 317 (5843): 1378–1381. Bibcode:2007Sci...317.1378T. doi:10.1126/science.1144066. PMID 17823350.
- ↑ Goehlich U.B., Tischlinger H., Chiappe L.M. (2006). "Juraventaor starki (Reptilia, Theropoda) ein nuer Raubdinosaurier aus dem Oberjura der Suedlichen Frankenalb (Sueddeutschland): Skelettanatomie und Wiechteilbefunde". Archaeopteryx. 24: 1–26.
- ↑ Chiappe, Luis M.; Göhlich, Ursula B. (2010). "Anatomy of Juravenator starki (Theropoda: Coelurosauria) from the Late Jurassic of Germany". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 258 (3): 257–296. doi:10.1127/0077-7749/2010/0125.
- ↑ Ji S., Ji Q., Lu J., Yuan C. (2007). "A new giant compsognathid dinosaur with long filamentous integuments from Lower Cretaceous of Northeastern China". Acta Geologica Sinica. 81 (1): 8–15.
- 1 2 Zhang, Fucheng; Zhou, Zhonghe; Xu, Xing; Wang, Xiaolin; Sullivan, Corwin (2008). "A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers". Nature. 455 (7216): 1105–1108. Bibcode:2008Natur.455.1105Z. doi:10.1038/nature07447. PMID 18948955.
- ↑ He T., Wang X.-L., Zhou Z.-H. (2008). "A new genus and species of caudipterid dinosaur from the Lower Cretaceous Jiufotang Formation of western Liaoning, China". Vertebrata PalAsiatica. 46 (3): 178–189.
- ↑ Xu, Xing; Zheng, Xiaoting; You, Hailu (2010). "Exceptional dinosaur fossils show ontogenetic development of early feathers". Nature. 464 (7293): 1338–1341. Bibcode:2010Natur.464.1338X. doi:10.1038/nature08965. PMID 20428169.
- ↑ Xu, X., Zhao, Q., Norell, M., Sullivan, C., Hone, D., Erickson, G., Wang, X., Han, F. and Guo, Y. (in press). "A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin." Chinese Science Bulletin, 6 pages, accepted 15 November 2008.
- 1 2 3 Zheng, Xiao-Ting; You, Hai-Lu; Xu, Xing; Dong, Zhi-Ming (2009). "An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures". Nature. 458 (7236): 333–336. Bibcode:2009Natur.458..333Z. doi:10.1038/nature07856. PMID 19295609.
- ↑ Xing Xu, Hailu You, Kai Du and Fenglu Han (28 July 2011). "An Archaeopteryx-like theropod from China and the origin of Avialae". Nature. 475 (7357): 465–470. doi:10.1038/nature10288. PMID 21796204.
- ↑ Xu, Xing; Wang, Kebai; Zhang, Ke; Ma, Qingyu; Xing, Lida; Sullivan, Corwin; Hu, Dongyu; Cheng, Shuqing; Wang, Shuo (2012). "A gigantic feathered dinosaur from the Lower Cretaceous of China". Nature. 484 (7392): 92–95. Bibcode:2012Natur.484...92X. doi:10.1038/nature10906. PMID 22481363.
- ↑ Rauhut, O. W. M.; Foth, C.; Tischlinger, H.; Norell, M. A. (2012). "Exceptionally preserved juvenile megalosauroid theropod dinosaur with filamentous integument from the Late Jurassic of Germany". Proceedings of the National Academy of Sciences. 109 (29): 11746–11751. Bibcode:2012PNAS..10911746R. doi:10.1073/pnas.1203238109. PMC 3406838. PMID 22753486.
- ↑ Darla K. Zelenitsky, François Therrien, Gregory M. Erickson, Christopher L. DeBuhr, Yoshitsugu Kobayashi, David A. Eberth and Frank Hadfield (2012). "Feathered Non-Avian Dinosaurs from North America Provide Insight into Wing Origins". Science. 338 (6106): 510–514. Bibcode:2012Sci...338..510Z. doi:10.1126/science.1225376. PMID 23112330.
- ↑ Ji Qiang, Lü Jun-Chang, Wei Xue-Fang, Wang Xu-Ri (2012). "A new oviraptorosaur from the Yixian Formation of Jianchang, Western Liaoning Province, China". Regional Geology of China (12): 2102–2107.
- ↑ Pascal Godefroit, Helena Demuynck, Gareth Dyke, Dongyu Hu, François Escuillié and Philippe Claeys (2013). "Reduced plumage and flight ability of a new Jurassic paravian theropod from China". Nature Communications. 4: Article number 1394. Bibcode:2013NatCo...4E1394G. doi:10.1038/ncomms2389. PMID 23340434.
- ↑ Hanyong Pu, Yoshitsugu Kobayashi, Junchang Lü, Li Xu, Yanhua Wu, Huali Chang, Jiming Zhang, Songhai Jia (2013). "An unusual basal therizinosaur dinosaur with an ornithischian dental arrangement from northeastern China". PLoS ONE. 8 (5): Article number e63423. Bibcode:2013PLoSO...863423P. doi:10.1371/journal.pone.0063423.
- ↑ Pascal Godefroit, Andrea Cau, Hu Dong-Yu, François Escuillié, Wu Wenhao, and Gareth Dyke (2013). "A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds". Nature. 498 (7454): 359–362. Bibcode:2013Natur.498..359G. doi:10.1038/nature12168. PMID 23719374.
- ↑ Gang Han, Luis M. Chiappe, Shu-An Ji, Michael Habib, Alan H. Turner, Anusuya Chinsamy, Xueling Liu, Lizhuo Han (2014). "A new raptorial dinosaur with exceptionally long feathering provides insights into dromaeosaurid flight performance". Nature Communications. 5: Article number 4382. Bibcode:2014NatCo...5E4382H. doi:10.1038/ncomms5382.
- ↑ Godefroit, Pascal; Sinitsa, Sofia M.; Dhouailly, Danielle; Bolotsky, Yuri L.; Sizov, Alexander V.; McNamara, Maria E.; Benton, Michael J.; Spagna, Paul (2014). "A Jurassic ornithischian dinosaur from Siberia with both feathers and scales". Science. 25 (345): 451–455. Bibcode:2014Sci...345..451G. doi:10.1126/science.1253351.
- 1 2 W. Scott Persons, IV, Philip J. Currie, and Mark A. Norell (2014). "Oviraptorosaur tail forms and functions". Acta Palaeontologica Polonica. doi:10.4202/app.2012.0093.
- ↑ Yuong-Nam Lee, Rinchen Barsbold, Philip J. Currie, Yoshitsugu Kobayashi, Hang-Jae Lee, Pascal Godefroit, François Escuillié, and Tsogtbaatar Chinzorig (2014). "Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus". Nature. 515 (7526): 257–260. Bibcode:2014Natur.515..257L. doi:10.1038/nature13874. PMID 25337880.
- ↑ Xu, X.; Zheng, X.; Sullivan, C.; Wang, X.; Xing, L.; Wang, Y.; Zhang, X.; O’Connor, J. K.; Zhang, F.; Pan, Y. (2015). "A bizarre Jurassic maniraptoran theropod with preserved evidence of membranous wings". Nature. 521 (7550): 70–3. Bibcode:2015Natur.521...70X. doi:10.1038/nature14423. PMID 25924069.
- ↑ Lü, Junchang; Brusatte, Stephen L. (2015). "A large, short-armed, winged dromaeosaurid (Dinosauria: Theropoda) from the Early Cretaceous of China and its implications for feather evolution". Scientific Reports. 5: 11775. doi:10.1038/srep11775. PMC 4504142. PMID 26181289.
- ↑ Robert A. DePalma, David A. Burnham, Larry D. Martin, Peter L. Larson, and Robert T. Bakker (2015). "The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation". Paleontological Contributions. 14: 1–15.
- ↑ Gregory F. Funston; Philip J. Currie (2016). "A new caenagnathid (Dinosauria: Oviraptorosauria) from the Horseshoe Canyon Formation of Alberta, Canada, and a reevaluation of the relationships of Caenagnathidae". Journal of Vertebrate Paleontology. Online edition (4): e1160910. doi:10.1080/02724634.2016.1160910.
- ↑ Czerkas, S.A., and Ji, Q. (2002). "A new rhamphorhynchoid with a headcrest and complex integumentary structures." Pp. 15-41 in: Czerkas, S.J. (Ed.). Feathered Dinosaurs and the Origin of Flight. Blanding, Utah: The Dinosaur Museum. ISBN 1-932075-01-1.
- ↑ Prum, R. & Brush A.H.; Brush (2002). "The evolutionary origin and diversification of feathers". The Quarterly Review of Biology. 77 (3): 261–295. doi:10.1086/341993. PMID 12365352.
- ↑ Prum, R (1999). "Development and evolutionary origin of feathers". Journal of Experimental Zoology. 285 (4): 291–306. doi:10.1002/(SICI)1097-010X(19991215)285:4<291::AID-JEZ1>3.0.CO;2-9. PMID 10578107.
- ↑ Norell, M. Xu, X. (2005) "The Varieties of Tyrannosaurs" Natural History Magazine May 2005.
- 1 2 Dimond, C. C.,R. J. Cabin and J. S. Brooks (2011). "Feathers, Dinosaurs, and Behavioral Cues: Defining the Visual Display Hypothesis for the Adaptive Function of Feathers in Non-Avian Theropods". BIOS. 82 (3): 58–63. doi:10.1893/011.082.0302.
- ↑ Sumida, S. S.; C. A. Brochu (2000). "Phylogenetic Context for the Origin of Feathers". American Zoologist. 40 (4): 485–503. doi:10.1093/icb/40.4.486.
- ↑ Lingham-Soliar, T. (2011). "The evolution of the feather: Sinosauropteryx, a colourful tail". Journal of Ornithology. 152 (3): 567–577. doi:10.1007/s10336-010-0620-y.
- ↑ Vinther, J.; D. E. G. Briggs; R. O. Prum & V. Saranathan (2008). "The colour of fossil feathers". Biology Letters. 4 (5): 522–525. doi:10.1098/rsbl.2008.0302. PMC 2610093. PMID 18611841.
- ↑ Zhang, F. C., S. L. Kearns, P. J. Orr, M. J. Benton, Z. H. Zhou, D. Johnson, X. Xu X. L. Wang (2010). "Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds". Nature. 463 (7284): 1075–1078. Bibcode:2010Natur.463.1075Z. doi:10.1038/nature08740. PMID 20107440.
- ↑ Zelenitsky, Darla K.; Therrien, Francois; Erickson, Gregory M.; DeBuhr, Christopher L.; Kobayashi, Yoshitsugu; Eberth, David A.; Hadfield, Frank (10/26/12). "Feathered Non-Avian Dinosaurs From North America Provide Insight into Wing Origins" Science 338 (510). doi:10.1126/science.1225376
- ↑ Li, Q. G.,K. Q. Gao,Q. J. Meng,M. D. Shawkey,L. D'Alba,R. Pei,M. Ellison,M. A. Norell and J. Vinther (2012). "Reconstruction of Microraptor and the Evolution of Iridescent Plumage". Science. 335 (6073): 1215–1219. Bibcode:2012Sci...335.1215L. doi:10.1126/science.1213780. PMID 22403389.
- ↑ Stevens, K.A. (2006). "Journal of Vertebrate Paleontology". 26: 11746–11751.
- ↑ Gould, Stephen J.; Vrba, Elisabeth S. (1982). "Exaptation: a missing term in the science of form" (PDF). Paleobiology. 8 (1): 4–15.
- ↑ Reichholf, J. H. (1996). "Die Feder, die Mauser und der Urspring der Vögel. Ein neure Sicht zur Evolution der Vögel". Archaeopteryx. 1427 (38).
- ↑ Bock, W. J. (2000). "Explanatory History of the Origin of Feathers". Am. Zool. 40 (4): 478–485. doi:10.1093/icb/40.4.478.
- ↑ Xu, X.; Zheng, X.; You, H. (2009). "A new feather type in a nonavian theropod and the early evolution of feathers". Proceedings of the National Academy of Sciences. 106 (3): 832–4. Bibcode:2009PNAS..106..832X. doi:10.1073/pnas.0810055106. PMC 2630069. PMID 19139401.
- ↑ "Ornithischian with complex feather-like structures found in Siberia (SVP abstract)".
- ↑ Zelenitsky, D. K.; Therrien, F.; Erickson, G. M.; Debuhr, C. L.; Kobayashi, Y.; Eberth, D. A.; Hadfield, F. (2012). "Feathered Non-Avian Dinosaurs from North America Provide Insight into Wing Origins". Science. 338 (6106): 510–514. Bibcode:2012Sci...338..510Z. doi:10.1126/science.1225376. PMID 23112330.
- ↑ Godefroit, Pascal; Cau, Andrea; Hu, Dong-Yu; Escuillié, François; Wu, Wenhao; Dyke, Gareth (2013). "A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds". Nature. 498 (7454): 359–362. Bibcode:2013Natur.498..359G. doi:10.1038/nature12168. PMID 23719374.
External links
- DinoBuzz, dinosaur-bird controversy explained, by UC Berkeley.