Endurance running hypothesis
The endurance running hypothesis is the hypothesis that the evolution of certain human characteristics can be explained as adaptations to long distance running.[1][2] The hypothesis suggests that endurance running played an important role for early hominins in obtaining food. Researchers have proposed that endurance running began as an adaptation for scavenging and later for persistence hunting.
Anatomical and physiological adaptations
Running vs. walking
Much research has been geared towards the mechanics of how bipedal walking has evolved in the genus Homo. However, little research has been conducted to examine how the specific adaptations for running emerged, and how they influenced human evolution.
The bit of research that has focused on human running provides much evidence for bodily function and structures that improve running only, and are not used in walking. This suggests that running was an adaptation, not that it came about as a byproduct of walking.
Running and walking incorporated different biomechanisms. Walking requires an "inverted pendulum" where the body's center of mass is shifted over the extended leg, to exchange potential and kinetic energy with each step.[2] Running involves a "mass spring" mechanism to exchange potential and kinetic energy, with the use of tendons and ligaments. Tendons and ligaments are elastic tissues that store energy. They are stretched and then release energy as they recoil. This mass spring mechanism becomes less energetically costly at faster speeds and is therefore more efficient than the inverted pendulum of walking mechanics when traveling at greater speeds.[2] Tendons and ligaments, however, don't provide these benefits in walking.
Although the mass spring mechanism can be more energetically favorable at higher speeds, it also results in an increase in ground reaction forces[2] and is less stable because there is more movement and pitching of the limbs and core of the body. Ground forces and body pitching movement is less of an issue in the walking gait, where the position of the body's center of mass varies less, making walking an inherently more stable gait. In response to the destabilization of the running gait, the human body appears to have evolved adaptations to increased stabilization, as well as for the mass-spring mechanism in general. These adaptations, described below, are all evidence for selection for endurance running.
Skeletal evidence
Many researchers compare the skeletal structures of early hominins such as Australopithecus to those of Homo in order to identify structural differences that may be significant to endurance running.
Nuchal ligament: Because the head is decoupled from the shoulders, early Homo needed a way to stabilize the head. The nuchal ligament is an important evolved feature in head stabilization. It starts at the midline of the occiput and connects to the upper trapezius. This ligament is also important in terms of archaeological findings, because it leaves a small indentation and ridge in the skull, allowing researchers to see if various species had a nuchal ligament. The ability to see traces of ligaments in archaeological findings is rare because they degrade quickly and often leave no trace. In the case of the nuchal ligament, a trace of its existence is left with the presence of the skull ridge. Because neither Australopithecus nor Pan had the skull ridge, it has been concluded that this feature is unique to Homo.[3]
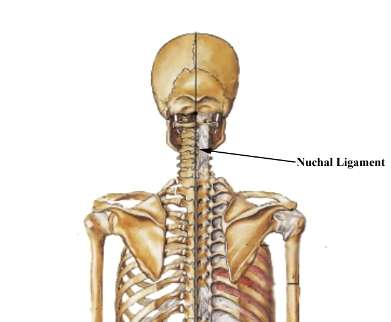
Shoulder and head stabilization: The human skeleton is different from early hominins as there is less of a connection between the pectoral girdle parts of the shoulders and upper back and head, which would be advantageous for climbing but would hinder the movements of the upper body needed to counter leg movement and therefore stabilize the body and head when running. This stabilization is unnecessary in walking.
Limb length and mass: Homo has longer legs relative to body mass, which helps to decrease energetic costs of running, as time in contact with the ground increases.[2] There is also a decrease in mass of distal parts of limbs of humans, which is known to decrease metabolic costs in endurance running, but has little effect on walking.[2] Additionally, the mass of the upper body limbs in Homo have decreased considerably, relative to total body mass, which is important to reduce effort of stabilizing the arms in running.
Joint surface: Humans have evolved to absorb great shock and force on the skeletal structure while running. The impact force on the body can reach up to 3-4 times body weight in endurance running, putting the skeletal structure under great stress. To reduce this stress humans have increased joint surfaces relative to body mass to spread force over larger surface areas, particularly in the lower body.[2] This adaptation, which allows humans to absorb great shock and force applied to the skeleton, is not seen in Australopithecine skeletal structures.
Plantar arch: The plantar arch in the human foot has an elastic spring function that generates energy for running but not walking.[2] Fossils of the australopithecine foot show only partial arch, suggesting less of a spring capacity. For the plantar arch spring mechanism to function fully, there must also be restricted rotation in the hind and front parts of the foot. This restriction comes from projected toe bone and compacted mid-foot joint structures in humans, which does not become present until Homo habilis.[2]
Calcaneal tuber and Achilles tendon: Studies have explored the calcaneal tuber, the posterior half of the calcaneus bone, as a correlate for Achilles tendon length and have found correlation between calcaneal tuber length and Achilles tendon length. Because shorter calcaneal tuber length leads to greater Achilles stretch, more kinetic energy is converted to elastic energy, translating into better overall running economy. Comparisons between Neanderthals and modern humans reveal that this adaptation was absent in Neanderthals, leading researchers to conclude that endurance running capabilities, including a well-developed Achilles tendon, did not evolve until at least the genus Homo.[4]
-

Calcaneus
Shorter toes: Human toes are straight and extremely short in relation to body size compared to other animals. In running, the toes support 50 to 75% of body mass in humans. Impulse and mechanical work increase in humans as toe length increases, showing that it is energetically favorable to have shorter toes. The costs of shorter toes are decreased gripping capabilities and power output. However, the efficiency benefits seemed to outweigh these costs, as the toes of A. afarensis remains were shorter than great apes, but 40% longer than modern humans, meaning that there is a trend toward shorter toes as the primate species moves away from tree-dwelling. This 40% increase in toe length would theoretically induce a flexor impulse 2.5 times that of modern humans, which would require twice as much mechanical work to stabilize.[5]
Stabilization
Semicircular canal: The semicircular canal, a series of three interconnected tubes within each ear, is important for sensing angular rotations of the head and thus plays a crucial role in maintaining balance and sensing and coordinating movement. Comparative studies have shown that animals with larger semicircular canals are able to sense a greater range of head movements and therefore have greater speed and agility. Evolutionarily, greatly reduced semicircular canal diameters are evident in Neanderthals but expanded in modern humans, suggesting that this adaptation was selected for in response to increased endurance running.[4]
Vestibulo-ocular reflexes (VOR's): VORs are enabled by muscles in the eye, which sense angular accelerations of the head and adjust eye movements to stabilize these images. This was an important adaptation for running because it allowed Homo to see more clearly during the rough pitching motion that occurs during running.[3]
Gluteals: The gluteus maximus in Homo erectus is significantly larger than that of Australopithecus. It is suited to stabilize the trunk while running, but gluteals of that size and strength are not necessary for walking.
Iliac Spine: Homo has expanded areas on the sacrum and posterior iliac spine for greater muscle attachment. These areas are used to stabilize the trunk and reduce body's forward pitch caused by running strides.
Increased efficiency
Thermoregulation
In addition to advances in skeletal structure and stabilization, adaptations that led to increased efficiency in dissipation of heat were instrumental in the evolution of endurance running in Homo.[1] The duration for which an animal can run is determined by its capacity to release more heat than is produced to avoid lethal temperatures.
- Evaporative Cooling: The majority of mammals, including humans, rely on evaporative cooling to maintain body temperature.
- Sweating vs. Panting: Most medium-to-large mammals rely on panting, while humans rely on sweating, to dissipate heat. Advantages to panting include cooler skin surface, little salt loss, and heat loss by forced convection instead of reliance on wind or other means of convection. On the other hand, sweating is advantageous in that evaporation occurs over a much larger surface area (the skin), and it is independent of respiration, thus is a much more flexible mode of cooling during intense activity such as running.
- Human Advantages: Because human sweat glands are under a higher level of neuronal control than those of other species, they allow for the excretion of more sweat per unit surface area than any other species. Heat dissipation of later hominins was also enhanced by the reduction in body hair.[1] By ridding themselves of an insulating fur coat, running humans are better able to dissipate the heat generated by exercise.
Respiration
In addition to improved thermoregulation, hominins have evolved an enhanced method of respiration consistent with the demands of running.
- Quadrupeds vs. Humans: Due to their orientation, respiration in quadrupedal mammals is affected by skeletal and muscular stresses generated through the motion of running. The bones and muscles of the chest cavity are not only responsible for shock absorption, but are also subjected to continuous compression and expansion during the running cycle. Because of this movement, quadrupeds are restricted to one breath per locomotor cycle, and thus must coordinate their running gait and respiration rate.[1] This tight coordination then translates into another restriction: a specific running speed that is most energetically favorable. The upright orientation of bipedal hominins, however, frees them from this respiration-gait restriction. Because their chest cavities are not directly compressed or involved in the motion of running, hominins are able to vary their breathing patterns with gait.[1] This flexibility in respiration rate and running gait contributes to hominins having a broader range of energetically favorable running speeds.
Storage and Utilization of Energy
During periods of prolonged exercise, animals are dependent on a combination of two sources of fuel: glycogen stored in the muscles and liver, and fat. Because glycogen is more easily oxidized than fat, it is depleted first. However, over longer periods of time, energy demands require that fat stores be utilized as fuel. This is true for all mammals, but hominins, and later modern humans, have an advantage of being able to alter their diet to meet these prolonged energy demands.
In addition to flexibility in the utilization of energy, hominins have evolved larger thyroid and adrenal glands which enable them to utilize the energy in carbohydrates and fatty acids more readily and efficiently. These organs are responsible for releasing hormones including epinephrine, norepinephrine, adrenocorticotrophic hormone (ACTH), glucagon, and thyroxine. Larger glands allows for greater production of these key hormones and ultimately, maximized utilization of stored fuel.[1]
Taken together, the flexibility in diet and the enhanced usage of fuel heightens the previously mentioned finding that, unlike quadrupeds, hominins do not have a single energetically optimal running speed. For quadrupeds, increasing running speed means increasing the demand for oxygen and fuel. Due to skeletal structure and bipedalism, hominins are free to run energetically over a broader range of speeds and gaits, while maintaining a constant energy consumption rate of approximately 4.1 MJ per 15 km. Thus their utilization of energy is greatly enhanced.[1]
Endurance running and scavenging
All of the aforementioned adaptations enabled Homo to scavenge for food more effectively. Endurance running could have been used as a means of gaining access to distant carcasses or food stores faster than other scavengers and/or carnivores. Scavenging may have taken one or both of two forms: Opportunistic scavenging and strategic scavenging.
Early Homo almost certainly scavenged opportunistically. Scavenging is considered opportunistic when one "come[s] across carcasses in the course of [their] daily foraging activities".[3]
Strategic scavenging involves a planned search for carcasses. This style of scavenging would have benefitted from endurance running much more than opportunistic scavenging. Strategic scavenging would have involved the use of long range cues, such as birds circling overhead. Endurance running would have been advantageous in this setting because it allowed hominins to reach the carcass quicker. Selection pressures would have been very high for strategic scavenging, because hominins were diurnal, while their major competitors (hyenas, lions, etc.) were not. Thus, they would have had to make sure to capitalize on daytime carcasses. Selection pressure also came from the weakness of Homo. Because they were very weak, they were unlikely to drive off any large competition at the carcass. This fact led to an even higher need for a way to reach the carcass before these competitors.[3]
Endurance running and persistence hunting
Persistence hunting is "a form of pursuit hunting in which humans use endurance running during the midday heat to drive animals into hyperthermia and exhaustion so they can easily be killed".[1][3] Many question persistence hunting's plausibility when bow and arrow and other technologies were so much more efficient. However, in the Early Stone Age (ESA), spears were only sharpened wood, and hominins had not begun using tools. The lack of spearheads or bows meant they could only hunt from very close range—between 6 and 10 meters.[6] Hominins thus must have developed a way to stab prey from close range without causing serious bodily harm to themselves. Persistence hunting makes killing an animal easier by first bringing it to exhaustion, so that it can no longer retaliate violently.
Persistence hunters work by hunting in the middle of the day, when it is hottest. Hunters choose a single target prey and chase it at a speed between its trot and gallop, which is extremely inefficient for the animal. The hunter then continues pursuing over a period of hours, during which he may lose sight of the animal. In this case, the hunter must use tracks and an understanding of the animal to continue the chase. The prey eventually overheats and becomes unable to continue fleeing. Homo, which does not overheat as quickly because of its superior thermoregulation capabilities, is then able to stab the prey while it is incapacitated and cannot attack.
Tracking
Due to the complexity of following a fleeing animal, tracking methods must have been a prerequisite for the use of endurance running in persistence hunting. Scientists posit that early tracking methods were developed in open, sparsely vegetated terrain such as the Kalahari Desert in southern Africa. This "systemic tracking" involves simply following the footprints of animals and was most likely used for tracking grassland species on soft terrain. Skeletal remains suggest that during the Middle Stone Age, hominins used systemic tracking to scavenge for medium-sized animals in vegetation cover, but for hunting antelope in more open grasslands. From the Middle Stone Age into the Later Stone Age, tracking methods developed into what is termed "speculative tracking". When tracks could not easily be found and followed, Homo predicted where tracks were most likely to be found and interpreted other signs to locate prey.[7] This advanced method of tracking allowed for the exploitation of prey in a variety of terrains, making endurance running for persistence hunting more plausible.
Examples of persistence hunters
Although exact dates and methods of persistence hunting are difficult to study, several recent accounts of persistence hunting have been recorded. Tribes in the Kalahari Desert in Botswana have been known to employ endurance running to scavenge and hunt prey. In the open country, the !Xo and /Gwi tribes run down slow-moving animals such as aardvark and porcupines, while during the hotter part of the day, they target animals such as eland, kudu, gemsbok, hartebeest, duiker, steenbok, cheetah, caracal, and African wildcats. In addition to these existing African tribes, it has been suggested that the Tarahumara people in Mexico and the Paiute people and Navajo in the American Southwest, used persistence hunting to capture prey including deer and pronghorn. The Aborigines in Australia are known to have hunted kangaroo in similar ways.[7] Due to the increased availability of weapons, nutrition, tracking devices, and motor vehicles, one may argue that persistence hunting is no longer an effective method of hunting animals for food. However, there are examples of the practice occurring in modern times: the !Xo and /Gwi in the central Kalahari, still practice persistence hunting and have developed very intelligent and advanced methods of doing so. Similarly, the Russian Lykov family that lived in isolation for 40 years also used persistence hunting due to a lack of weapons.[8]



Seasonal differences
In particular, these two tribes maximize the efficiency of persistence hunting by targeting specific species during different seasons. In the rainy season, prime targets include steenbok, duiker, and gemsbok, as wet sand opens their hooves and stiffens their joints. Hunting in the early rainy season is particularly advantageous because dry leaves form "rocks" in the animals' stomachs, resulting in diarrhea. Stiff joints and suboptimal digestion make the prey weaker and more available targets. In contrast, in the dry season, hunters run down kudu, eland, and red hartebeest because these species tire more easily in the loose sand. Hunters say that the best time to persistent hunt is near the end of the dry season when animals are poorly nourished and therefore more easily run to exhaustion.[7] By targeting the most vulnerable prey during each season, the hunters maximize the advantages of endurance running.
Conditions/parameters
1. Persistence hunting must be performed during the day when it is hot, so that the animal will overheat.
2. Homo must have been able to track the animal, as they would have lost sight of it during the chase.
3. Such a long hunt requires high amounts of dietary sources of water, salt, and glycogen.
4. Although the success rate of recorded persistence hunts is very high (approximately 50%[3] ), unsuccessful hunts are very costly. Therefore, there would have had to be a social system in which individuals share food, so unsuccessful hunters could borrow food from others when necessary.
Criticisms
While there is evidence supporting selection on human morphology to improve endurance running ability, there is some dispute over whether the ecological benefits of scavenging and persistence hunting foraging behaviors were the driving force behind this development.
The majority of the arguments opposing persistence hunting and scavenging behaviors are linked to the fact that the paleohabitat and paleoecology of early Homo were not conducive to these behaviors. It is thought that the earliest members of Homo lived in African savanna-woodlands.[9] This environment consisted of open grassland, as well as parts with dense vegetation - an intermediate between forest and open savannas. The presence of such tree covering would reduce visibility and so require tracking skills. This causes problems for the hypothesis of persistence hunting and running to aid scavenging.
Against persistence hunting
Ungulates are known from archaeological evidence to have been the main prey of the early Homo, and given their great speed, they would have easily been able to outrun early hominins. Ungulate speed, coupled with the variable visibility of the savanna–woodland, meant that hunting by endurance running required the ability to track prey. Pickering and Bunn argue that tracking is part of a sophisticated cognitive skill set that early hominins would not have had, and that even if they were following a trail of blood left by an injured ungulate—which may have been in their cognitive capacity—the ability to craft penetrating projectile technology was absent in early hominins.[9]
It has been suggested that modern hunters in Africa do not use persistence hunting as a foraging method, and most often give up a chase where the trail they were following ends in vegetation.[9] The rare groups of hunters who do occasionally participate in persistence hunting are able to do so because of the extremely hot and open environments. In these groups, a full day of rest and recovery is required after a hunt, indicating the great toll persistence hunts take on the body and making them rare undertakings.[9]
Finally, in critique of Liebenberg's research on modern day persistence hunting, it was revealed that the majority of the hunts initiated were prompted for filming rather than spontaneous, and that few of these hunts were successful. The hunts that were successful involved external factors such as the hunters being able to stop and refill water bottles.[9]
A response to these criticisms has been formulated by Lieberman et al.[10]
Against scavenging
The proposed benefit of endurance running in scavenging is the ability of early hominins to outcompete other scavengers in reaching food sources.[9] However paleoanthropological studies suggest that the savanna-woodland habitat caused a very low competition environment. Due to low visibility, carcasses were not easily located by mammalian carnivores, resulting in less competition.
References
- 1 2 3 4 5 6 7 8 Carrier, D.R.; et al. (Aug–Oct 1984). "The Energetic Paradox of Human Running and Hominid Evolution". Current Anthropology. 25 (4): 483–495. doi:10.1086/203165. JSTOR 2742907.
- 1 2 3 4 5 6 7 8 9 Bramble, Dennis; Lieberman, Daniel (November 2004). "Endurance running and the evolution of Homo". Nature. 432: 345–52. doi:10.1038/nature03052. PMID 15549097.
- 1 2 3 4 5 6 Lieberman, Daniel; Bramble, Dennis; Raichlen, David; Shea, John (October 2006). "Brains, Brawn, and the Evolution of Human Endurance Running Capabilities". Contributions from the Third Stony Brook Human Evolution Symposium and Workshop: 77–92.
- 1 2 Raichlen, D.A.; Armstrong, H; Lieberman, D.E. (26 January 2011). "Calcaneus length determines running economy: Implications for endurance running performance in modern humans and Neandertals.". Journal of Human Evolution. 60: 299–308. doi:10.1016/j.jhevol.2010.11.002.
- ↑ Rolian, Campbell; Lieberman, Daniel; Hamill, Joseph; Scott, John; Werbel, William (November 2008). "Walking, running and the evolution of short toes in humans". The Journal of Experimental Biology: 212, 713–721.
- ↑ Churchill, S.E. (1993). "Weapon technology, prey size selection and hunting methods in modern hunter gatherers: implications for hunting in the Paleolithic and Mesolithic". Archeological Papers of the American Anthropological Association (4): 11–24.
- 1 2 3 Liebenberg, Louis (December 2006). "Persistence Hunting by Modern Hunter-Gatherers". Current Anthropology. 47 (6): 1017–1026. doi:10.1086/508695. JSTOR 10.1086/508695.
- ↑ http://www.smithsonianmag.com/history-archaeology/For-40-Years-This-Russian-Family-Was-Cut-Off-From-Human-Contact-Unaware-of-World-War-II-188843001.html
- 1 2 3 4 5 6 Pickering, Travis Rayne; Bunn, Henry (October 2007). "The endurance running hypothesis and hunting and scavenging in savanna-woodlands". Journal of Human Evolution. 53 (4): 434–438. doi:10.1016/j.jhevol.2007.01.012.
- ↑ "The evolution of endurance running and the tyranny of ethnography: A reply to Pickering and Bunn (2007)" (PDF).