List of diseases of the honey bee
Diseases of the honey bee or abnormal hive conditions include:
Pests and parasites
Varroa mites
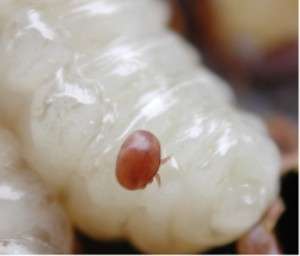
Varroa destructor and Varroa jacobsoni are parasitic mites that feed on the bodily fluids of adult, pupal and larval bees. Varroa mites can be seen with the naked eye as a small red or brown spot on the bee's thorax. Varroa mites are carriers for a virus that is particularly damaging to the bees. Bees infected with this virus during their development will often have visibly deformed wings.
Varroa mites have led to the virtual elimination of feral bee colonies in many areas, and are a major problem for kept bees in apiaries. Some feral populations are now recovering—it appears they have been naturally selected for Varroa resistance.
Varroa mites were first discovered in Southeast Asia in about 1904, but are now present on all continents except Australia. They were discovered in the United States in 1987, in New Zealand in 2000, and in Devon, United Kingdom in 1992.
These mites are generally not a problem for a strongly growing hive. When the hive population growth is reduced in preparation for winter or due to poor late summer forage, the mite population growth can overtake that of the bees and can then destroy the hive. Often a colony will simply abscond (leave as in a swarm, but leaving no population behind) under such conditions.
Varroa in combination with deformed wing virus and bacteria have been theoretically implicated in colony collapse disorder.
Treatment
A variety of treatments are currently marketed or practiced to attempt to control these mites. The treatments are generally segregated into chemical and mechanical controls.
Common chemical controls include "hard" chemicals such as Amitraz (marketed as Apivar [1]), fluvalinate (marketed as Apistan), and coumaphos (marketed as CheckMite). "Soft" chemical controls include thymol (marketed as ApiLife-VAR and Apiguard), sucrose octanoate esters (marketed as Sucrocide), oxalic acid and formic acid (sold in gel packs as Mite-Away,[2] but also used in other formulations). According to the U.S. Environmental Protection Agency, when used in beehives as directed, these treatments kill a large proportion of the mites while not substantially disrupting bee behavior or life span. Use of chemical controls is generally regulated and varies from country to country. With few exceptions, they are not intended for use during production of marketable honey. [3]
Common mechanical controls generally rely on disruption of some aspect of the mites' lifecycle. These controls are generally intended not to eliminate all mites, but merely to maintain the infestation at a level which the colony can tolerate. Examples of mechanical controls include drone brood sacrifice (varroa mites are preferentially attracted to the drone brood), powdered sugar dusting (which encourages cleaning behavior and dislodges some mites), screened bottom boards (so any dislodged mites fall through the bottom and away from the colony), brood interruption and, perhaps, downsizing of the brood cell size. A device called the varroa mite control entrance (VMCE) is under development as of 2008. The VMCE works in conjunction with a screened bottom board, by dislodging varroa mites from bees as they enter and exit a hive.[4]
Acarine (tracheal) mites
Acarapis woodi is a small parasitic mite that infests the airways of the honey bee. The first known infestation of the mites occurred in the British Isles in the early 20th century. First observed on the Isle of Wight in 1904,[5] the mystery illness known as Isle of Wight Disease was not identified as being caused by a parasite until 1921. It quickly spread to the rest of Great Britain. It was regarded as having wiped out the entire native bee population of the British Isles (although later genetic studies have found remnants that did survive) and it dealt a devastating blow to British beekeeping. Brother Adam at the Buckfast Abbey developed a resistant hybrid bee known as the Buckfast bee, which is now available worldwide to combat acarine disease.
Diagnosis for tracheal mites generally involves the dissection and microscopic examination of a sample of bees from the hive.
Acarine mites, formerly known as tracheal mites, are believed to have entered the US in 1984, from Mexico.
Mature female acarine mites leave the bee's airway and climb out on a hair of the bee, where they wait until they can transfer to a young bee. Once on the new bee, they move into the airways and begin laying eggs.
Treatment
Acarine mites are commonly controlled with grease patties (typically made from one part vegetable shortening mixed with three to four parts powdered sugar) placed on the top bars of the hive. The bees come to eat the sugar and pick up traces of shortening, which disrupts the mite's ability to identify a young bee. Some of the mites waiting to transfer to a new host remain on the original host. Others transfer to a random bee—a proportion of which will die of other causes before the mite can reproduce.
Menthol, either allowed to vaporize from crystal form or mixed into the grease patties, is also often used to treat acarine mites.
Nosema disease
Nosema apis is a microsporidian that invades the intestinal tracts of adult bees and causes nosema disease, also known as nosemosis. Nosema infection is also associated with black queen cell virus. It is normally only a problem when the bees cannot leave the hive to eliminate waste (for example, during an extended cold spell in winter or when the hives are enclosed in a wintering barn). When the bees are unable to void (cleansing flights), they can develop dysentery.
Nosema disease is treated by increasing the ventilation through the hive. Some beekeepers treat hives with antibiotics such as fumagillin.
Nosemosis can also be prevented or minimized by removing much of the honey from the beehive, then feeding the bees on sugar water in the late fall. Sugar water made from refined sugar has lower ash content than flower nectar, reducing the risk of dysentery. Refined sugar, however, contains fewer nutrients than natural honey, which causes some controversy among beekeepers.
In 1996, a similar type of organism to N. apis was discovered on the Asian honey bee Apis cerana and subsequently named Nosema ceranae. This parasite apparently also infects the western honey bee.[6]
Exposure to corn pollen containing genes for Bacillus thuringiensis (Bt) production may weaken the bees' defense against Nosema.[7] In relation to feeding a group of bees with Bt corn pollen and a control group with non-Bt corn pollen: "in the first year, the bee colonies happened to be infested with parasites (microsporidia). This infestation led to a reduction in the number of bees and subsequently to reduced broods in the Bt-fed colonies, as well as in the colonies fed on Bt toxin-free pollen. The trial was then discontinued at an early stage. This effect was significantly more marked in the Bt-fed colonies. (The significant differences indicate an interaction of toxin and pathogen on the epithelial cells of the honeybee intestine. The underlying mechanism which causes this effect is unknown.)"
This study should be interpreted with caution given that no repetition of the experiment nor any attempt to find confounding factors was made. In addition, Bt toxin and transgenic Bt pollen showed no acute toxicity to any of the life stages of the bees examined, even when the Bt toxin was fed at concentrations 100 times that found in transgenic Bt pollen from maize.
Small hive beetle

Aethina tumida is a small, dark-colored beetle that lives in beehives. Originally from Africa, the first discovery of small hive beetles in the Western Hemisphere was made in St. Lucie County, Florida, in 1998. The next year, a specimen that had been collected from Charleston, South Carolina, in 1996 was identified, and is believed to be the index case for the United States.[8] By December 1999, small hive beetles were reported in Iowa, Maine, Massachusetts, Minnesota, New Jersey, Ohio, Pennsylvania, Texas, and Wisconsin, and it was found in California by 2006.
The lifecycle of this beetle includes pupation in the ground outside of the hive. Controls to prevent ants from climbing into the hive are believed to also be effective against the hive beetle. Several beekeepers are experimenting with the use of diatomaceous earth around the hive as a way to disrupt the beetle's lifecycle. The diatoms abrade the insects' surfaces, causing them to dehydrate and die.
Treatment
Several pesticides are currently used against the small hive beetle. The chemical fipronil (marketed as Combat Roach Gel [9]) is commonly applied inside the corrugations of a piece of cardboard. Standard corrugations are large enough that a small hive beetle can enter the cardboard through the end, but small enough that honey bees cannot enter (thus are kept away from the pesticide). Alternative controls such as oil-based top-bar traps are also available, but they have had very little commercial success.
Wax moths

Galleria mellonella (greater wax moths) do not attack the bees directly, but feed on the wax used by the bees to build their honeycomb. Their full development to adults requires access to used brood comb or brood cell cleanings—these contain protein essential for the larval development, in the form of brood cocoons. The destruction of the comb will spill or contaminate stored honey and may kill bee larvae.
When honey supers are stored for the winter in a mild climate, or in heated storage, the wax moth larvae can destroy portions of the comb, though they will not fully develop. Damaged comb may be scraped out and replaced by the bees. Wax moth larvae and eggs are killed by freezing, so storage in unheated sheds or barns in higher latitudes is the only control necessary.
Because wax moths cannot survive a cold winter, they are usually not a problem for beekeepers in the northern U.S. or Canada, unless they survive winter in heated storage, or are brought from the south by purchase or migration of beekeepers. They thrive and spread most rapidly with temperatures above 30 °C (90 °F), so some areas with only occasional days that are hot rarely have a problem with wax moths, unless the colony is already weak due to stress from other factors.
Control and treatment
A strong hive generally needs no treatment to control wax moths; the bees themselves kill and clean out the moth larvae and webs. Wax moth larvae may fully develop in cell cleanings when such cleanings accumulate thickly where they are not accessible to the bees.
Wax moth development in comb is generally not a problem with top bar hives, as unused combs are usually left in the hive during the winter. Since this type of hive is not used in severe wintering conditions, the bees are able to patrol and inspect the unused comb.
Wax moths can be controlled in stored comb by application of the aizawai variety of Bacillus thuringiensis spores by spraying. It is a very effective biological control and has an excellent safety record.
Wax moths can be controlled chemically with paradichlorobenzene (moth crystals or urinal disks). If chemical methods are used, the combs must be well-aired for several days before use. The use of naphthalene (mothballs) is discouraged because it accumulates in the wax, which can kill bees or contaminate honey stores. Control of wax moths by other means includes the freezing of the comb for at least 24 hours.
Tropilaelaps
Tropilaelaps clareae and T. mercedesae are considered threats to honeybees. Although they are not currently found in the US or Canada, these mites have the potential to inflict serious damage to colonies due to their rapid reproduction inside the hive.
Bacterial diseases
| Appearance of brood comb | Age of dead brood | Color of dead brood | Consistency of dead brood | Odor of dead brood | Scale characteristics | Infectious agent |
|---|---|---|---|---|---|---|
| Sealed brood, discolored, sunken, or punctured cappings | Usually older sealed larvae or young pupae lying lengthwise in cells | Dull white, becoming light brown, coffee brown to dark brown, or almost black | Soft, becoming sticky to ropy | Slightly to pronounced putrid odor | Lies uniformly flat on lower side of cell, adheres tightly to cell wall, fine, thread-like tongue of dead may be present, head lies flat, black in color | American foulbrood |
| Unsealed brood, some sealed brood in advanced cases with discolored, sunken or punctured cappings | Usually young unsealed larvae, occasionally older sealed larvae, typically in coiled stage | Dull white, becoming yellowish white to brown, dark brown, or almost black | Watery, rarely sticky or ropy, granular | Slightly to penetrating sour | Usually twisted in cell, does not adhere to cell wall, rubbery, black in color | European foulbrood |
American foulbrood
American foulbrood (AFB), caused by the spore-forming Paenibacillus larvae[11] (formerly classified as Bacillus larvae and Paenibacillus larvae ssp larvae/pulvifaciens), is the most widespread and destructive of the bee brood diseases. P. larvae is a rod-shaped bacterium. Larvae up to three days old become infected by ingesting spores present in their food. Young larvae less than 24 hours old are most susceptible to infection. Spores germinate in the gut of the larva and the vegetative bacteria begin to grow, taking nourishment from the larva. Spores will not germinate in larvae over three days old. Infected larvae normally die after their cell is sealed. The vegetative form of the bacterium will die, but not before it produces many millions of spores. American foulbrood spores are extremely resistant to desiccation and can remain viable for more than 40 years in honey and beekeeping equipment. Each dead larva may contain as many as 100 million spores. This disease only affects the bee larvae, but is highly infectious and deadly to bee brood. Infected larvae darken and die.
As with European foulbrood, research has been conducted using the 'Shook Swarm'[12] method to control American foulbrood, "the advantage being that chemicals are not used".
European foulbrood
Melissococcus plutonius is a bacterium that infects the midgut of bee larvae. European foulbrood is considered less serious than American foulbrood. M. plutonius is not a spore-forming bacterium, but bacterial cells can survive for several months on wax foundation. Symptoms include dead and dying larvae which can appear curled upwards, brown or yellow, melted or deflated with tracheal tubes more apparent, or dried out and rubbery.[13]
European foulbrood is often considered a "stress" disease — dangerous only if the colony is already under stress for other reasons. An otherwise healthy colony can usually survive European foulbrood. Chemical treatment with oxytetracycline hydrochloride may control an outbreak of the disease, but honey from treated colonies could have chemical residues from the treatment. The "Shook Swarm"[14] technique of bee husbandry can also effectively control the disease, with the advantage of avoiding the use of chemicals. Prophylactic treatments are not recommended as they lead to resistant bacteria.
Fungal diseases
Chalkbrood
_L.S._Olive_%26_Spiltoir_1324048.jpg)
Ascosphaera apis is a fungal disease that infests the gut of the larva. The fungus competes with the larva for food, ultimately causing it to starve. The fungus then goes on to consume the rest of the larva's body, causing it to appear white and 'chalky'.
Chalkbrood is most commonly visible during wet springs. Hives with chalkbrood can generally be recovered by increasing the ventilation through the hive.
Stonebrood
Stonebrood is a fungal disease caused by Aspergillus fumigatus, Aspergillus flavus, and Aspergillus niger. It causes mummification of the brood of a honey bee colony. The fungi are common soil inhabitants and are also pathogenic to other insects, birds, and mammals. The disease is difficult to identify in the early stages of infection. The spores of the different species have different colours and can also cause respiratory damage to humans and other animals. When bee larvae take in spores, they may hatch in the gut, growing rapidly to form a collar-like ring near the larval heads. After death, the larvae turn black and become difficult to crush, hence the name stonebrood. Eventually, the fungus erupts from the integument of the larvae and forms a false skin. In this stage, the larvae are covered with powdery fungal spores. Worker bees clean out the infected brood and the hive may recover depending on factors such as the strength of the colony, the level of infection, and hygienic habits of the strain of bees (variation in the trait occurs among different subspecies/races).
Viral diseases
Cripaviridae
Chronic bee paralysis virus
- Syndrome 1 Abnormal trembling of the wings and body occurs. The bees cannot fly, and often crawl on the ground and up plant stems. In some cases, the crawling bees can be found in large numbers (1000+). The bees huddle together on the top of the cluster or on the top bars of the hive. They may have bloated abdomens due to distension of the honey sac. The wings are partially spread or dislocated.
- Syndrome 2 Affected bees are able to fly, but are almost hairless. They appear dark or black and look smaller. They have a relatively broad abdomen. They are often nibbled by older bees in the colony and this may be the cause of the hairlessness. They are hindered at the entrance to the hive by the guard bees. A few days after infection, trembling begins. They then become flightless and soon die.[15][16]
In 2008, the chronic bee paralysis virus was reported for the first time in Formica rufa and another species of ant, Camponotus vagus.[17]
Dicistroviridae
Acute bee paralysis virus
Acute bee paralysis virus[18] is considered to be a common infective agent of bees. It belongs to the family Dicistroviridae,[19] as does the Israel acute paralysis virus, Kashmir bee virus, and the black queen cell virus. It is frequently detected in apparently healthy colonies. Apparently, this virus plays a role in cases of sudden collapse of honey bee colonies infested with the parasitic mite Varroa destructor.[20]
Israeli acute paralysis virus
A related virus[19] described in 2004 is known as the Israeli acute paralysis virus (IAPV).[21] The virus is named after the place where it was first identified—its place of origin is unknown. IAPV has been suggested as a marker associated with colony collapse disorder.[22][23]
Kashmir bee virus
Kashmir bee virus[24] is related to the preceding viruses.[19] Recently discovered, it is currently only positively identifiable by a laboratory test. Little is known about it yet.[25]
Black queen cell virus
Black queen cell virus[26] causes the queen larva to turn black and die. It is thought to be associated with Nosema.[27]
Cloudy wing virus
Cloudy wing virus is a little-studied, small, icosahedral virus commonly found in honey bees, especially in collapsing colonies infested by Varroa destructor, providing circumstantial evidence that the mite may act as a vector.[28][29][30]
Sacbrood virus
A picornavirus-like causes sacbrood disease.[31][32] Affected larvae change from pearly white to gray and finally black. Death occurs when the larvae are upright, just before pupation. Consequently, affected larvae are usually found in capped cells. Head development of diseased larvae is typically retarded. The head region is usually darker than the rest of the body and may lean toward the center of the cell. When affected larvae are carefully removed from their cells, they appear to be a sac filled with water. Typically, the scales are brittle but easy to remove. Sacbrood-diseased larvae have no characteristic odor.[10][33]
Iflaviridae
Deformed wing virus
Deformed wing virus (DWV) is the causative agent of the wing deformities and other body malformations typically seen in honeybee colonies that are heavily infested with the parasitic mite Varroa destructor.[34] DWV is part of a complex of closely related virus strains/species that also includes Kakugo virus, Varroa destructor virus 1[35] and Egypt bee virus. This deformity can clearly be seen on the honeybee's wings in the image. The deformities are produced almost exclusively due to DWV transmission by V. destructor when it parasitizes pupae. Bees infected as adults remain symptom-free, although they do display behavioral changes and have reduced life expectancy. Deformed bees are rapidly expelled from the colony, leading to a gradual loss of adult bees for colony maintenance. If this loss is excessive and can no longer be compensated by the emergence of healthy bees, the colony rapidly dwindles and dies.
Kakugo virus
Kakugo virus is an iflavirus infecting bees; varroa mites may mediate its prevalence.[36]
Iridoviridae
Invertebrate iridescent virus type 6
Applying proteomics-based pathogen screening tools in 2010, researchers announced they had identified a co-infection of an iridovirus;[37] specifically invertebrate iridescent virus type 6 (IIV-6) and Nosema ceranae in all CCD colonies sampled.[38] On the basis of this research, the New York Times reported the colony collapse mystery solved, quoting researcher Dr. Bromenshenk, a co-author of the study, "[The virus and fungus] are both present in all these collapsed colonies."[39][40] Evidence for this association, however, remains minimal[41] and several authors have disputed the original methodology used to associate CCD with IIV-6.[42][43]
Secoviridae
Tobacco ringspot virus
The RNA virus tobacco ringspot virus, primarily a plant pathogen, can infect honeybees through infected pollen.[44]
Lake Sinai virus
Lake Sinai virus (LSV) genomes were assembled and three main domains were discovered: Orf1, RNA-dependent RNA polymerase and capsid protein sequences. LSV1, LSV2, LSV3, LSV4, LSV5, and LSV6 are described.[45] LSV are detected in bees, mites and pollen, only actively replicates in honey bees and mason bees (Osmia cornuta) and not in Varroa mites.[46]
Dysentery
Dysentery is a condition resulting from a combination of long periods of inability to make cleansing flights (generally due to cold weather) and food stores which contain a high proportion of indigestible matter. As a bee's gut becomes engorged with feces that cannot be voided in flight as preferred by the bees, the bee voids within the hive. When enough bees do this, the hive population rapidly collapses and death of the colony results. Dark honeys and honeydews have greater quantities of indigestible matter.
Occasional warm days in winter are critical for honey bee survival; dysentery problems increase in likelihood during periods of more than two or three weeks with temperatures below 50 °F (10 °C). When cleansing flights are few, bees will often be forced out at times when the temperature is barely adequate for their wing muscles to function, and large quantities of bees may be seen dead in the snow around the hives. Colonies found dead in spring from dysentery will have feces smeared over the frames and other hive parts.
In very cold areas of North America and Europe, where honey bees are kept in ventilated buildings during the coldest part of winter, no cleansing flights are possible; under such circumstances, it is common for beekeepers to remove all honey from the hives and replace it with sugar water or high fructose corn syrup, which have nearly no indigestible matter.
Chilled brood
Chilled brood is not actually a disease, but can be a result of mistreatment of the bees by the beekeeper. It also can be caused by a pesticide hit that primarily kills off the adult population, or by a sudden drop in temperature during rapid spring buildup. The brood must be kept warm at all times; nurse bees will cluster over the brood to keep it at the right temperature. When a beekeeper opens the hive (to inspect, remove honey, check the queen, or just to look) and prevents the nurse bees from clustering on the frame for too long, the brood can become chilled, deforming or even killing some of the bees.
Pesticide losses
Honey bees are susceptible to many of the chemicals used for agricultural spraying of other insects and pests. Many pesticides are known to be toxic to bees. Because the bees forage up to several miles from the hive, they may fly into areas actively being sprayed by farmers or they may collect pollen from contaminated flowers.
Carbamate pesticides, such as carbaryl, can be especially pernicious since toxicity can take as long as two days to become evident, allowing infected pollen to be returned and distributed throughout the colony. Organophosphates and other insecticides are also known to kill honey bee clusters in treated areas.
Pesticide losses may be relatively easy to identify (large and sudden numbers of dead bees in front of the hive) or quite difficult, especially if the loss results from a gradual accumulation of pesticide brought in by the foraging bees. Quick-acting pesticides may deprive the hive of its foragers, dropping them in the field before they can return home.
Insecticides that are toxic to bees have label directions that protect the bees from poisoning as they forage. To comply with the label, applicators must know where and when bees forage in the application area, and the length of residual activity of the pesticide.
Some pesticide authorities recommend, and some jurisdictions require, that notice of spraying be sent to all known beekeepers in the area, so they can seal the entrances to their hives and keep the bees inside until the pesticide has had a chance to disperse. This, however, does not solve all problems associated with spraying and the label instructions should be followed regardless of doing this. Sealing honey bees from flight on hot days can kill bees. Beekeeper notification does not offer any protection to bees, if the beekeeper cannot access them, or to wild native or feral honey bees. Thus, beekeeper notification as the sole protection procedure does not really protect all the pollinators of the area, and is, in effect, a circumventing of the label requirements. Pesticide losses are a major factor in pollinator decline.
Colony collapse disorder
Colony collapse disorder (CCD) is a poorly understood phenomenon in which worker bees from a beehive or western honey bee colony abruptly disappear. CCD was originally discovered in Florida by David Hackenberg in western honey bee colonies in late 2006.[47]
European beekeepers observed a similar phenomenon in Belgium, France, the Netherlands, Greece, Italy, Portugal, and Spain,[48] and initial reports have also come in from Switzerland and Germany, albeit to a lesser degree.[49] Possible cases of CCD have also been reported in Taiwan since April 2007.[50]
Initial hypotheses were wildly different, including environmental change-related stresses,[51] malnutrition, pathogens (i.e., disease[52] including Israel acute paralysis virus[53][54]), mites, or the class of pesticides known as neonicotinoids, which include imidacloprid, clothianidin, and thiamethoxam. Unproven and less credible theories include radiation from cellular phones or other man-made devices,[55] and genetically modified crops with pest-control characteristics.[56] In 2010, US researchers announced they had identified a co-infection of invertebrate iridescent virus type 6 (IIV-6) and Nosema ceranae in all CCD colonies sampled.[57]
References
- ↑ "Home". apivar.net.
- ↑ "NOD - Nature's Own Design". miteaway.com.
- ↑ Office of Pesticide Programs; US Environmental Protection Agency Formic Acid (214900) Fact Sheet (2005)
- ↑ http://www.scienceinafrica.co.za/2008/october/debugging.htm
- ↑ Anderson, John; Rennie, John (1916). Observations and experiments bearing on the "Island of Wight" disease. Edinburgh: R. Grant & Son.
- ↑ Ritter, Wolfgang Nosema ceranae Albert Ludwigs University of Freiburg
- ↑ "Effects of Bt maize pollen on the honeybee". 2005-10-12. Archived from the original on 2007-01-26. Retrieved 2007-03-21.
- ↑ Neumann P, Elzen PJ (2004). "The biology of the small hive beetle (Aethina tumida, Coleoptera: Nitidulidae): Gaps in our knowledge of an invasive species". Apidologie. 35 (3): 229–247. doi:10.1051/apido:2004010.
- ↑ "Combat Source Kill Max Gel Kills Large and Small Roaches". combatbugs.com.
- 1 2 Shimanuki, Hachiro; Knox, David A. Diagnosis of Honey Bee Diseases USDA Archived 9 December 2006 at the Wayback Machine.
- ↑ Genersch, E; Forsgren, E; Pentikäinen, J; Ashiralieva, A; Rauch, S; Kilwinski, J; Fries, I (March 2006). "Reclassification of Paenibacillus larvae subsp. pulvifaciens and Paenibacillus larvae subsp. larvae as Paenibacillus larvae without subspecies differentiation.". International Journal of Systematic and Evolutionary Microbiology. 56 (Pt 3): 501–11. doi:10.1099/ijs.0.63928-0. PMID 16514018.
- ↑ Controlling American Foulbrood in Honeybees By Shook Swarm Method
- ↑ "European Foulbrood: A Bacterial Disease Affecting Honey Bee Brood". extension.org.
- ↑ "Shook Swarm and OTC Antibiotics for European Foulbrood Control". extension.org.
- ↑ Ribière M, Faucon J, Pépin M (2000). "Detection of chronic honey bee (Apis mellifera L.) paralysis virus infection: application to a field survey" (PDF). Apidologie. 31 (5): 567–77. doi:10.1051/apido:2000147.
- ↑ "Chronic Bee Paralysis Virus". globalnet.co.uk.
- ↑ Detection of Chronic bee paralysis virus (CBPV) genome and its replicative RNA form in various hosts and possible ways of spread.
- ↑ (TaxID 92444)
- ↑ Bakonyi T, Grabensteiner E, Kolodziejek J, et al. (December 2002). "Phylogenetic analysis of acute bee paralysis virus strains". Appl. Environ. Microbiol. 68 (12): 6446–50. doi:10.1128/AEM.68.12.6446-6450.2002. PMC 134446
 . PMID 12450876.
. PMID 12450876. - ↑ (TaxID 294365)
- ↑ Fox, Maggie (2007-09-06). "New virus may be killing bees". Reuters. Retrieved 2007-09-06.
- ↑ Humberto F. Boncristiani; et al. "In vitro infection of pupae with IAPV suggests disturbance of transcriptional homeostasis in honey bees (Apis mellifera)". PLOS ONE. 8: e73429. doi:10.1371/journal.pone.0073429.
- ↑ (TaxID 68876)
- ↑ Kashmir Bee Virus Apiculture Factsheet #230, Ministry of Agriculture and Lands, Government of British Columbia, Jul 2004, accessed Jan 2007
- ↑ (TaxID 92395)
- ↑ "Multiple Virus Infections in the Honey Bee and Genome Divergence of Honey Bee Viruses". usda.gov.
- ↑ Carreck, Norman L; Ball, Brenda V; Martin, Stephen J (2010). "The epidemiology of cloudy wing virus infections in honey bee colonies in the UK". Journal of Apicultural Research. 49 (1): 66–71. doi:10.3896/ibra.1.49.1.09.
- ↑ Gliński, Z.; Jarosz, J. (2001). "Infection and immunity in the honey bee Apis mellifera". Apiacta. 36 (1): 12–24.
- ↑ Cloudy Wing Virus Universal Protein Resource (UniProt); European Bioinformatics Institute (EBI), the Swiss Institute of Bioinformatics (SIB) and the Protein Information Resource (PIR)
- ↑ Wu, CY; Lo, CF; Huang, CJ; Yu, HT; Wang, CH (2002). "The complete genome sequence of Perina nuda picorna-like virus, an insect-infecting RNA virus with a genome organization similar to that of the mammalian picornaviruses". Virology. 294 (2): 312–323. doi:10.1006/viro.2001.1344.
- ↑ Grabensteiner, E; Ritter, W; Carter, MJ; Davison, S; Pechhacker, H; Kolodziejek, J; Boecking, O; Derakhshifar, I; Moosbeckhofer, R; Licek, E; Nowotny, N (2001). "Sacbrood Virus of the Honeybee (Apis mellifera): Rapid Identification and Phylogenetic Analysis Using Reverse Transcription-PCR". Clinical and Diagnostic Laboratory Immunology. 8 (1): 93–104. doi:10.1128/CDLI.8.1.93-104.2001.
- ↑ Grabensteiner, Elvira; Ritter, Wolfgang; Carter, Michael J.; Davison, Sean; Pechhacker, Hermann; Kolodziejek, Jolanta; Boecking, Otto; Derakhshifar, Irmgard; Moosbeckhofer, Rudolf; Licek, Elisabeth; Nowotny, Norbert (2001). "Sacbrood Virus of the Honeybee (Apis mellifera): Rapid Identification and Phylogenetic Analysis Using Reverse Transcription-PCR". Clin Diagn Lab Immunol. 8 (1): 93–104. doi:10.1128/CDLI.8.1.93-104.2001. PMC 96016. PMID 11139201.
- ↑ de Miranda, Joachim R.; Genersch, Elke (2010). "Deformed wing virus". Journal of Invertebrate Pathology. 103: S48–S61. doi:10.1016/j.jip.2009.06.012. PMID 19909976.
- ↑ Juliette R. Ongus, Els C. Roode, Cornelis W. A. Pleij, Just M. Vlak and Monique M. van Oers The 5' non-translated region of Varroa destructor virus 1 (genus Iflavirus): structure prediction and IRES activity in Lymantria dispar cell J Gen Virol 87 (2006), 3397-3407; doi:10.1099/vir.0.82122-0
- ↑ Tomoko Fujiyuki, Seii Ohka, Hideaki Takeuchi, Masato Ono, Akio Nomoto, and Takeo Kubo Prevalence and Phylogeny of Kakugo Virus, a Novel Insect Picorna-Like Virus That Infects the Honeybee (Apis mellifera L.), under Various Colony Conditions Journal of Virology, December 2006, p. 11528-11538, Vol. 80, No. 23
- ↑ Williams, Trevor Iridoviridae Instituto de Ecología AC (INECOL)Mexico
- ↑ Leal, Walter S.; Bromenshenk, Jerry J.; Henderson, Colin B.; Wick, Charles H.; Stanford, Michael F.; Zulich, Alan W.; Jabbour, Rabih E.; Deshpande, Samir V.; McCubbin, Patrick E. (2010). Leal, Walter S., ed. "Iridovirus and Microsporidian Linked to Honey Bee Colony Decline". PLoS ONE. 5 (10): e13181. doi:10.1371/journal.pone.0013181. PMC 2950847. PMID 20949138.
- ↑ Kirk Johnson (2010-10-06). "Scientists and Soldiers Solve a Bee Mystery". The New York Times.
- ↑ Invertebrate iridescent virus type 6 (IIV-6) Universal Protein Resource (UniProt); European Bioinformatics Institute (EBI), the Swiss Institute of Bioinformatics (SIB) and the Protein Information Resource (PIR).
- ↑ Tokarz, Rafal; Cadhla Firth; Craig Street; Diana Cox-Foster; W. Ian Lipkin (2011). "Lack of evidence for an Association between Iridovirus and Colony Collapse Disorder". PLoS ONE. 6 (6): e21844. doi:10.1371/journal.pone.0021844.
- ↑ Foster, Leonard (2011). "Interpretation of data underlying the link between colony collapse disorder (CCD) and an invertebrate iridescent virus.". Molecular & Cellular Proteomics. 10 (3).
- ↑ Knudsen, Giselle; Robert Chalkley (2011). "The Effect of Using an Inappropriate Protein Database for Proteomic Data Analysis". PLoS ONE. 6 (6): e20873. doi:10.1371/journal.pone.0020873. PMC 3114852. PMID 21695130.
- ↑ Li, J. L.; Cornman, R. S.; Evans, J. D.; Pettis, J. S.; Zhao, Y.; Murphy, C.; Peng, W. J.; Wu, J.; Hamilton, M.; Boncristiani, H. F.; Zhou, L.; Hammond, J.; Chen, Y. P. (21 January 2014). "Systemic Spread and Propagation of a Plant-Pathogenic Virus in European Honeybees, Apis mellifera". MBio. 5 (1): e00898–13–e00898–13. doi:10.1128/mBio.00898-13.
- ↑ Daughenbaugh, Katie F.; et al. "Honey Bee Infecting Lake Sinai Viruses". Viruses. 7: 3285–3309. doi:10.3390/v7062772.
- ↑ Ravoet, Jorgen; et al. "Genome sequence heterogeneity of Lake Sinai Virus found in honey bees and Orf1/RdRP-based polymorphisms in a single host". Virus Research. 201: 67–72. doi:10.1016/j.virusres.2015.02.019.
- ↑ "Honey Bee Die-Off Alarms Beekeepers, Crop Growers and Researchers". Penn State University College of Agricultural Sciences. 2007-01-29.
- ↑ Gaëlle Dupont, Les abeilles malades de l'homme, Le Monde, 29 August 2007 (French)
- ↑ Petra Steinberger (2007-03-12). "Das spurlose Sterben" (in German). sueddeutsche.de.
- ↑ Paul Molga, La mort des abeilles met la planète en danger, Les Echos, 20 August 2007 (French)
- ↑ Amy Sahba (2007-03-29). "The mysterious deaths of the honeybees". CNN Money. Retrieved 2007-04-04.
- ↑ "Colony Collapse Disorder Working Group".
- ↑ JR Minkel (2007-09-07). "Mysterious Honeybee Disappearance Linked to Rare Virus". Science News. Scientific American. Retrieved 2007-09-07.
- ↑ Andrew C. Refkin (2007-09-07). "Virus Is Seen as Suspect in Death of Honeybees". The New York Times. Retrieved 2007-09-07.
- ↑ Geoffrey Lean & Harriet Shawcross (2007-04-15). "Are mobile phones wiping out our bees?". The Independent. Retrieved 2007-12-10.
- ↑ "GE and bee Colony Collapse Disorder -- science needed!". 2005-03-21. Retrieved 2007-03-23.
- ↑ Leal, Walter S.; Bromenshenk, Jerry J.; Henderson, Colin B.; Wick, Charles H.; Stanford, Michael F.; Zulich, Alan W.; Jabbour, Rabih E.; Deshpande, Samir V.; McCubbin, Patrick E. (2010). Leal, Walter S., ed. "Iridovirus and Microsporidian Linked to Honey Bee Colony Decline". PLoS ONE. 5 (10): e13181. doi:10.1371/journal.pone.0013181. PMC 2950847. PMID 20949138.
Further reading
- Canadian Honey Council Essential Oils for Varroa, Tracheal, AFB Control
- Sammataro, Diana; et al. The Beekeeper's Handbook
- Morse, Roger (editor) The ABC and XYZ of Beekeeping
External links
- Beekeeping page at the University of Georgia, with a large section on Honey Bee Disorders
- Apiculture Factsheets at the British Columbia Ministry of Agriculture and Lands
- BeeBase at the Defra Food and Environment Research Agency in the UK
- Kohala.net Diseases and Afflictions of Honey Bees
- Beediseases Honey bee diseases website by Dr. Guido Cordoni.