DEAD box
| DEAD/DEAH box helicase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identifiers | |||||||||
| Symbol | DEAD | ||||||||
| Pfam | PF00270 | ||||||||
| Pfam clan | CL0023 | ||||||||
| InterPro | IPR011545 | ||||||||
| PROSITE | PDOC00039 | ||||||||
| SCOP | 1qva | ||||||||
| SUPERFAMILY | 1qva | ||||||||
| CDD | cd00268 | ||||||||
| |||||||||
DEAD box proteins are involved in an assortment of metabolic processes that typically involve RNAs, but in some cases also other nucleic acids.[2] They are highly conserved in nine motifs and can be found in most prokaryotes and eukaryotes, but not all. Many organisms, including humans, contain DEAD-box helicases, which are involved in RNA metabolism.[3]
DEAD Box Family
DEAD box proteins were first brought to attention in the late 1980s in a study that looked at a group of NTP binding sites that were similar in sequence to the eIF4A RNA helicase sequence.[4] The results of this study showed that these proteins (p68,SrmB, MSS116, vasa, PL10, mammalian eIF4A, yeast eIF4A) involved in RNA metabolism had several common elements.[5] There were nine common sequences found to be conserved amongst the studied species, which is an important criterion of the DEAD box family.[5] The nine conserved motifs are as follows, Q-motif, motif 1, motif 1a, motif 1b, motif II, motif III, motif IV, motif V, and motif VI, as shown in the figure. Motif II is also known as the Walker B motif and contains the amino acid sequence D-E-A-D (asp-glu-ala-asp), which gave this family of proteins the name “DEAD box”.[5] Motif 1, motif II, the Q motif, and motif VI are all needed for ATP binding and hydrolysis, while motifs, 1a, 1b, III, IV, and V may be involved in intramolecular rearrangements and RNA interaction.[6]

Related Families
The DEAH and SKI families have had proteins that have been identified to be related to the DEAD box family.[7][8][9] These two relatives have a few particularly unique motifs that are conserved within their own family. DEAD box, DEAH, and the SKI families are all referred to as DExD/H proteins.[10] They are all quite distinct from one another and there is not one protein that belongs to more than one of these families. It is thought that each family has a specific role in RNA metabolism, for example both DEAD box and DEAH box proteins NTPase activities become stimulated by RNA, but DEAD box proteins use ATP and DEAH does not.[6]
Biological Functions
DEAD box proteins are considered to be RNA helicases and many have been found to be required in cellular processes such as pre-mRNA processing and rearranging of ribonucleoproteins (RNP) complexes.[11]
Pre-mRNA Splicing
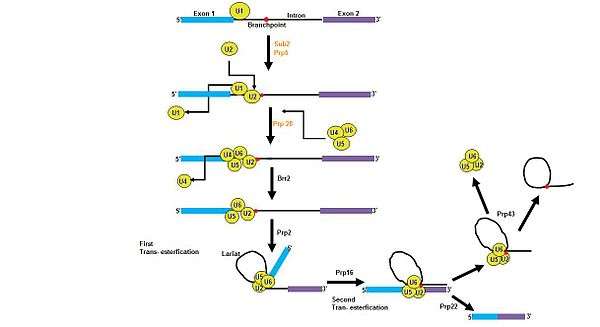
Pre-mRNA splicing requires rearrangements of five large RNP complexes, which are snRNPs U1, U2, U4, U5, and U6. DEAD box proteins are helicases that perform unwinding in an energy dependent approach and are able to perform these snRNP rearrangements in a quick and efficient manner.[12] There are three DEAD box proteins in the yeast system, Sub2, Prp28, and Prp5, and have been proven to be required for in vivo splicing.[12] Prp5 has been shown to assist in a conformational rearrangement of U2 snRNA, which makes the branch point recognition sequence of U2 available to bind the branch point sequence.[13] Prp28 may have a role in recognizing the 5’ splice site and does not display RNA helicase activity, suggesting that other factors must be present in order to activate Prp28.[14] DExD/H proteins have also been found to be required components in pre- mRNA splicing, in particular the DEAH proteins, Prp2, Prp16, Prp22, Prp43, and Brr213.[15] As shown in the figure, DEAD box proteins are needed in the initial steps of spliceosome formation, while DEAH box proteins are indirectly required for the transesterifications, release of the mRNA, and recycling of the spliceosome complex9.
Translation Initiation
The eIF4A translation initiation factor was the first DEAD box protein found to have a RNA dependent ATPase activity. It has been proposed that this abundant protein helps in unwinding the secondary structure in the 5’-untranslated region.[16] This can inhibit the scanning process of the small ribosomal subunit, if not unwound.[16] Ded1 is another DEAD box protein that is also needed for translation initiation, but its exact role in this process is still obscure.[17] Vasa, a DEAD box protein highly related to Ded1 plays a part in translation initiation by interacting with eukaryotic initiation factor 2 (eIF2).[18]
See also
- DEAD/DEAH box helicase
- Ski 2 helicase
- Walker A motif
- RNA helicase
- DDX3X
References
- ↑ Johnson, E. R.; McKay, D. B. (1999). "Crystallographic structure of the amino terminal domain of yeast initiation factor 4A, a representative DEAD-box RNA helicase". RNA. 5 (12): 1526–1534. doi:10.1017/S1355838299991410. PMC 1369875
 . PMID 10606264.
. PMID 10606264. - ↑ Takashi Kikuma; Masaya Ohtsu; Takahiko Utsugi; Shoko Koga; Kohji Okuhara; Toshihiko Eki; Fumihiro Fujimori; Yasufumi Murakami (March 2004). "Dbp9p, a Member of the DEAD Box Protein Family, Exhibits DNA Helicase Activity". J. Biol. Chem. 279 (20): 20692–20698. doi:10.1074/jbc.M400231200. PMID 15028736.
- ↑ Heung LJ, Del Poeta M (March 2005). "Unlocking the DEAD-box: a key to cryptococcal virulence?". J. Clin. Invest. 115 (3): 593–5. doi:10.1172/JCI24508. PMC 1052016. PMID 15765144.
- ↑ Gorbalenya AE, Koonin EV, Donchenko AP, Blinov VM (June 1989). "Two related superfamilies of putative helicases involved in replication, recombination, repair and expression of DNA and RNA genomes". Nucleic Acids Res. 17 (12): 4713–30. doi:10.1093/nar/17.12.4713. PMC 318027. PMID 2546125.
- 1 2 3 Linder, P.; Lasko, P. F.; Ashburner, M.; Leroy, P.; Nielsen, P. J.; Nishi, K.; Schnier, J.; Slonimski, P. P. (1989). "Birth of the D-E-A-D box". Nature. 337 (6203): 121–122. Bibcode:1989Natur.337..121L. doi:10.1038/337121a0. PMID 2563148.
- 1 2 Tanner NK, Cordin O, Banroques J, Doère M, Linder P (January 2003). "The Q motif: a newly identified motif in DEAD box helicases may regulate ATP binding and hydrolysis". Mol. Cell. 11 (1): 127–38. doi:10.1016/S1097-2765(03)00006-6. PMID 12535527.
- ↑ Tanaka N, Schwer B (July 2005). "Characterization of the NTPase, RNA-binding, and RNA helicase activities of the DEAH-box splicing factor Prp22". Biochemistry. 44 (28): 9795–803. doi:10.1021/bi050407m. PMID 16008364.
- ↑ Xu J, Wu H, Zhang C, Cao Y, Wang L, Zeng L, Ye X, Wu Q, Dai J, Xie Y, Mao Y (2002). "Identification of a novel human DDX40gene, a new member of the DEAH-box protein family". J. Hum. Genet. 47 (12): 681–3. doi:10.1007/s100380200104. PMID 12522690.
- ↑ Wang L, Lewis MS, Johnson AW (August 2005). "Domain interactions within the Ski2/3/8 complex and between the Ski complex and Ski7p". RNA. 11 (8): 1291–302. doi:10.1261/rna.2060405. PMC 1370812. PMID 16043509.
- ↑ de la Cruz J, Kressler D, Linder P (May 1999). "Unwinding RNA in Saccharomyces cerevisiae: DEAD-box proteins and related families". Trends Biochem. Sci. 24 (5): 192–8. doi:10.1016/S0968-0004(99)01376-6. PMID 10322435.
- ↑ Staley JP, Guthrie C (February 1998). "Mechanical devices of the spliceosome: motors, clocks, springs, and things". Cell. 92 (3): 315–26. doi:10.1016/S0092-8674(00)80925-3. PMID 9476892.
- 1 2 Linder P (2006). "Dead-box proteins: a family affair—active and passive players in RNP-remodeling". Nucleic Acids Res. 34 (15): 4168–80. doi:10.1093/nar/gkl468. PMC 1616962. PMID 16936318.
- ↑ Ghetti A, Company M, Abelson J (April 1995). "Specificity of Prp24 binding to RNA: a role for Prp24 in the dynamic interaction of U4 and U6 snRNAs". RNA. 1 (2): 132–45. PMC 1369067. PMID 7585243.
- ↑ Strauss EJ, Guthrie C (August 1994). "PRP28, a 'DEAD-box' protein, is required for the first step of mRNA splicing in vitro". Nucleic Acids Res. 22 (15): 3187–93. doi:10.1093/nar/22.15.3187. PMC 310295. PMID 7520570.
- ↑ Silverman E, Edwalds-Gilbert G, Lin RJ (July 2003). "DExD/H-box proteins and their partners: helping RNA helicases unwind". Gene. 312: 1–16. doi:10.1016/S0378-1119(03)00626-7. PMID 12909336.
- 1 2 Sonenberg N (1988). "Cap-binding proteins of eukaryotic messenger RNA: functions in initiation and control of translation". Prog. Nucleic Acid Res. Mol. Biol. Progress in Nucleic Acid Research and Molecular Biology. 35: 173–207. doi:10.1016/S0079-6603(08)60614-5. ISBN 978-0-12-540035-0. PMID 3065823.
- ↑ Berthelot K, Muldoon M, Rajkowitsch L, Hughes J, McCarthy JE (February 2004). "Dynamics and processivity of 40S ribosome scanning on mRNA in yeast". Mol. Microbiol. 51 (4): 987–1001. doi:10.1046/j.1365-2958.2003.03898.x. PMID 14763975.
- ↑ Carrera P, Johnstone O, Nakamura A, Casanova J, Jäckle H, Lasko P (January 2000). "VASA mediates translation through interaction with a Drosophila yIF2 homolog". Mol. Cell. 5 (1): 181–7. doi:10.1016/S1097-2765(00)80414-1. PMID 10678180.