Cynodont
| Cynodonts Temporal range: Late Permian-Holocene, 260–0 Ma | |
|---|---|
 | |
| Fossil of Belesodon magnificus in the Staatliches Museum für Naturkunde Stuttgart | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Order: | Therapsida |
| Clade: | Eutheriodontia |
| Suborder: | Cynodontia Owen, 1861 |
| Families | |
|
See text | |
The cynodonts ("dog teeth"), in the clade Cynodontia, are therapsids that first appeared in the Late Permian (approximately 260 Ma). The group includes modern mammals (including humans) as well as their extinct ancestors and close relatives. Nonmammalian cynodonts spread throughout southern Gondwana and are represented by fossils from South America, Africa, India, and Antarctica. In the northern continents, fossils have been found in eastern North America as well as in Belgium and northwestern France. Cynodontia is one of the most diverse groups of therapsids.
Taxonomy
Richard Owen named Cynodontia in 1861, which he assigned to Anomodontia as a family.[1] Robert Broom (1913) reranked Cynodonia as an infraorder, since retained by others, including Colbert and Kitching (1977), Carroll (1988), Gauthier et al. (1989), and Rubidge and Cristian Sidor (2001).[2] Olson (1966) assigned Cynodontia to Theriodonta, Colbert and Kitching (1977) to Theriodontia, and Rubridge and Sidor (2001) to Eutheriodontia. William King Gregory (1910), Broom (1913), Carroll (1988), Gauthier et al. (1989), Hopson and Kitching (2001) and Botha et al. (2007) all considered Cynodontia as belonging to Therapsida. Botha et al. (2007) seems to have followed Owen (1861), but without specifying taxonomic rank.[3][4]
Evolutionary history
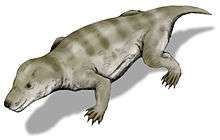
Together with the extinct gorgonopsians and the therocephalians, the cynodonts themselves are part of a group of therapsids called theriodonts.
The oldest and the most basal cynodont yet found is Charassognathus (Late Permian). Other basal cynodonts were the procynosuchids, a family that includes Procynosuchus and Dvinia. Cynodonts were among the few groups of synapsids that survived the Permian–Triassic extinction event and had a slow recovery after the extinction.
The most derived cynodonts are found within the clade Eucynodontia, which also contains the members of Mammalia. Representative genera of nonmammalian cynodonts include the large carnivorous cynognathids, the equally large herbivorous traversodonts, and the small mammal-like tritylodontids and ictidosaurs. The presence of respiratory turbinates suggests a rapid metabolism and possibly endothermy.
During their evolution, the number of cynodont jaw bones reduced. This move towards a single bone for the mandible paved the way for other bones in the jaw, the articular and angular, to migrate to the cranium, where they function as parts of the mammalian hearing system.
Cynodonts also developed a secondary palate in the roof of the mouth. This caused air flow from the nostrils to travel to a position in the back of the mouth instead of directly through it, allowing cynodonts to chew and breathe at the same time. This characteristic is present in all mammals.
Characteristics

Early cynodonts have many of the skeletal characteristics of mammals. The teeth were fully differentiated and the braincase bulged at the back of the head. Outside of some crown-group mammals (notably the therians), all cynodonts probably laid eggs. The temporal fenestrae were much larger than those of their ancestors, and the widening of the zygomatic arch in a more mammal-like skull would have allowed for more robust jaw musculature. They also have the secondary palate that other primitive therapsids lacked, except the therocephalians, who were the closest relatives of cynodonts. The dentary was the largest bone in their lower jaw.
The cynodonts probably had some form of warm-blooded metabolism. This has led to many reconstructions of cynodonts as having fur. Being endothermic they may have needed it for thermoregulation, but fossil evidence of their fur (or lack thereof) has been elusive. Modern mammals have Harderian glands secreting lipids to coat their fur, but the telltale imprint of this structure is only found from the primitive mammal Morganucodon and onwards.[5] Nonetheless, recent studies on Permian synapsid coprolites show that more basal therapsids had fur,[6] and at any rate fur was already present in Mammaliaformes such as Castorocauda and Megaconus.
Marks in the upper and lower jaw of cynodonts has been interpreted as channels that supplied blood vessels and nerves to whiskers.[7][8] Whiskers may have been typical of cynodontia as a whole, or have evolved in this group.
Derived cynodonts developed epipubic bones. These served to strengthen the torso and support abdominal and hindlimb musculature, aiding them in the development of an erect gait, but at the expense of prolonged pregnancy, forcing these animals to give birth to larval young as in modern monotremes and marsupials. Only placentals, and perhaps Megazostrodon and Erythrotherium, would lose these.[9][10]
Phylogeny



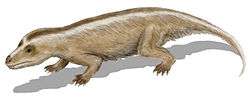

Below is a cladogram from Ruta, Botha-Brink, Mitchell and Benton (2013) showing one hypothesis of cynodont relationships:[11]
| Cynodontia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
| → Cynognathia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
| → Probainognathia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
See also
References
- ↑ Classification of R. Owen 1861.
- ↑ Classification of B. S. Rubidge and C. A. Sidor 2001
- ↑ R. Broom. 1913. A revision of the reptiles of the Karroo. Annals of the South African Museum 7(6):361-366
- ↑ S. H. Haughton and A. S. Brink. 1954. A bibliographical list of Reptilia from the Karroo Beds of South Africa. Palaeontologia Africana 2:1-187
- ↑ Ruben, J.A.; Jones, T.D. (2000). "Selective Factors Associated with the Origin of Fur and Feathers". American Zoologist. 40 (4): 585–596. doi:10.1093/icb/40.4.585.
- ↑ Microbiota and food residues including possible evidence of pre-mammalian hair in Upper Permian coprolites from Russia Piotr Bajdek1, Martin Qvarnström2, Krzysztof Owocki3, Tomasz Sulej3, Andrey G. Sennikov4,5, Valeriy K. Golubev4,5 andGrzegorz Niedźwiedzki2 Article first published online: 25 NOV 2015 DOI: 10.1111/let.12156
- ↑ Brink, A.S. (1955). "A study on the skeleton of Diademodon". Palaeontologia Africana. 3: 3–39.
- ↑ Kemp, T.S. (1982). Mammal-like reptiles and the origin of mammals. London: Academic Press. p. 363. ISBN 0-12-404120-5.
- ↑ Michael L. Power,Jay Schulkin. The Evolution Of The Human Placenta. pp. 68–.
- ↑ Jason A. Lillegraven, Zofia Kielan-Jaworowska, William A. Clemens, Mesozoic Mammals: The First Two-Thirds of Mammalian History, University of California Press, 17/12/1979 - 321
- ↑ Ruta, M.; Botha-Brink, J.; Mitchell, S. A.; Benton, M. J. (2013). "The radiation of cynodonts and the ground plan of mammalian morphological diversity". Proceedings of the Royal Society B: Biological Sciences. 280 (1769): 20131865. doi:10.1098/rspb.2013.1865.
Further reading
- Hopson, J.A.; Kitching, J.W. (2001). "A probainognathian cynodont from South Africa and the phylogeny of non-mammalian cynodonts". Bull. Mus. Comp. Zool. 156: 5–35.
- Davis, Dwight (1961). "Origin of the Mammalian Feeding Mechanism". Am. Zoologist, 1:229-234.
External links
- Palaeos cynodonts
- Phylogeny of Theriodonts and Cynodonts
- Bennett and Ruben 1986. The Metabolic and Thermoregulatory Status of Therapsids
- BBC cynodonts