Anomalocaris
| Anomalocaris Temporal range: Early to mid Cambrian: Chengjiang–Burgess shale | |
|---|---|
 | |
| Image of the first complete Anomalocaris fossil found, residing in the Royal Ontario Museum | |
| Scientific classification | |
| Kingdom: | Animalia |
| Class: | †Dinocaridida |
| Order: | †Radiodonta |
| Family: | †Anomalocarididae |
| Genus: | †Anomalocaris Whiteaves 1892 |
| Species | |
| |
| Synonyms | |
|
Defunct species:[1]
| |
Anomalocaris ("abnormal shrimp") is an extinct genus of anomalocaridid, a family of animals thought to be closely related to ancestral arthropods. The first fossils of Anomalocaris were discovered in the Ogygopsis Shale by Joseph Frederick Whiteaves, with more examples found by Charles Doolittle Walcott in the famed Burgess Shale.[2] Originally several fossilized parts discovered separately (the mouth, feeding appendages and tail) were thought to be three separate creatures, a misapprehension corrected by Harry B. Whittington and Derek Briggs in a 1985 journal article.[2][3]

Anatomy

Anomalocaris is thought to have been a predator. It propelled itself through the water by undulating the flexible lobes on the sides of its body.[4] Each lobe sloped below the one more posterior to it,[5] and this overlapping allowed the lobes on each side of the body to act as a single "fin", maximizing the swimming efficiency.[4] The construction of a remote-controlled model showed this mode of swimming to be intrinsically stable,[6] implying that Anomalocaris would not have needed a complex brain to manage balance while swimming. The body was widest between the third and fifth lobe and narrowed towards the tail; it had at least 11 lobes in total.[5] It is difficult to distinguish lobes near the tail, making an accurate count difficult.[5]
Anomalocaris had a large head, a single pair of large, compound eyes on stalks comprising approximately 16,000 lenses,[7][8] and an unusual disk-like mouth. The mouth was composed of 32 overlapping plates, four large and 28 small, resembling a pineapple ring with the center replaced by a series of serrated prongs.[2] The mouth could constrict to crush prey, but never completely close, and the tooth-like prongs continued down the walls of the gullet.[9] Two large 'arms' (up to 18 centimeters in length when extended[9]) with barb-like spikes were in front of the mouth.[3] The tail was large and fan-shaped, and along with undulations of the lobes, was probably used to propel the creature through Cambrian waters.[2][4][10] Stacked lamella of what were probably gills attached to the top of each lobe.
For the time in which it lived, Anomalocaris was gigantic, up to one meter long.[2]
Discovery
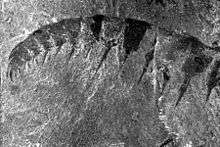
Anomalocaris has been misidentified several times, in part due to its makeup of a mixture of mineralized and unmineralized body parts; the mouth and feeding appendage was considerably harder and more easily fossilized than the delicate body.[9] Its name originates from a description of a detached 'arm', described by Joseph Frederick Whiteaves in 1892 as a separate crustacean-like creature due to its resemblance to the tail of a lobster or shrimp.[9] The first fossilized anomalocaridid mouth was discovered by Charles Doolittle Walcott, who mistook it for a jellyfish and placed it in the genus Peytoia. Walcott also discovered a second feeding appendage, but failed to realize the similarities to Whiteaves' discovery and instead identified it as feeding appendage or tail of the extinct Sidneyia.[9] The body was discovered separately and classified as a sponge in the genus Laggania; a mouth was found with the body, but was interpreted by its discoverer Simon Conway Morris as an unrelated Peytoia that had through happenstance settled and been preserved with Laggania. Later, while clearing what he thought was an unrelated specimen, Harry B. Whittington removed a layer of covering stone to discover the unequivocally connected arm thought to be a shrimp tail and mouth thought to be a jellyfish.[2][9] Whittington linked the two species, but it took several more years for researchers to realize that the continuously juxtaposed Peytoia, Laggania and feeding appendage actually represented a single, enormous creature.[9] Because Peytoia was named first, it became the correct name for the entire animal. The original feeding arm, however, came from a larger species distinct from Peytoia and "Laggania", which retains the name Anomalocaris.[11]
Stephen Jay Gould cites Anomalocaris as one of the fossilized extinct species he believed to be evidence of a much more diverse set of phyla that existed in the Cambrian Period[9] (discussed in his book Wonderful Life), a conclusion disputed by other paleontologists.[2]
In 2011, six fossils of compound eyes dated to the Cambrian period (515 million years ago) were recovered from an archaeological dig at Emu Bay on Kangaroo Island, Australia. The eyes were the first found that belonged to Anomalocaris, proving that Anomalocaris was indeed an arthropod as had been suspected. The find also indicated that advanced arthropod eyes able to "tell friend or foe and detect features of the environment" had evolved very early, before the evolution of jointed legs or hardened exoskeletons. The eyes were 30 times as powerful as those of trilobites, long thought to have had the most advanced eyes of any contemporary species. With 16,000 lenses, the resolution of the 3 centimetres (1.2 in) wide eyes would have been rivalled only by that of the modern dragonfly, which has 28,000 lenses in each eye.[12][13][14]
Ecology

Anomalocaris had a cosmopolitan distribution in Cambrian seas, and has been found from early to mid Cambrian deposits from Canada, China, Utah and Australia, to name but a few.[6][15][16][17]
A long-standing view holds that Anomalocaris fed on hard-bodied animals, including trilobites; while its mid-gut glands strongly suggest a predatory lifestyle, its ability to penetrate mineralised shells has, however, been questioned in recent years.[18] Some Cambrian trilobites have been found with round or W-shaped "bite" marks, which were identified as being the same shape as the mouthparts of Anomalocaris.[16] Stronger evidence that Anomalocaris ate trilobites comes from fossilised faecal pellets, which contain trilobite parts and are so large that the anomalocarids are the only known organism from that period large enough to have produced them.[16] However, since Anomalocaris lacks any mineralised tissue, it seemed unlikely that it would be able to penetrate the hard, calcified shell of trilobites.[16] Rather, the coprolites may have been produced by a different organism, such as a trilobite of the genus Redlichia.[19]

At most one species, A. canadensis, may have been capable of feeding on organisms with hard exoskeletons, due to the short, robust spines on its appendages.[19] However, the proposal that A. canadensis was durophagous is also not uncontested: studies of mouthparts associated with A. canadensis suggested that they are in fact not tetraradial like those of Peytoia, but have three major axes and a small central opening, which is more suggestive of suction feeding instead of durophagy.[20]
Other species of Anomalocaris likely preyed on soft-bodied animals. Recent phylogenetic analyses have also found "A." briggsi to be closer to the filter-feeding anomalocaridids (including Tamisiocaris) than Anomalocaris, which suggests that "A." briggsi is likely not a species of Anomalocaris and may have been a filter-feeder as well.[21][22]
One suggested possibility is that anomalocaridids fed by grabbing one end of their prey in their jaws while using their appendages to quickly rock the other end of the animal back and forth. This produced stresses that exploited the weaknesses of arthropod cuticles, causing the prey's exoskeleton to rupture and allowing the predator to access its innards.[16] This behaviour was originally thought to have provided an evolutionary pressure for trilobites to roll up, to avoid being flexed until they snapped.[16]
However, the lack of wear on anomalocaridid mouthparts suggests they did not come into regular contact with mineralised trilobite shells, and computer modelling of anomalocaridid mouthparts suggests they were possibly better suited to feeding on smaller, soft-bodied organisms by suction, since they would have experienced structural failure if they were used against the armour of trilobites.[18][23]
See also
- 8564 Anomalocaris, an asteroid, named after this animal
- Anomalocarididae, the group that Anomalocaris belongs to
- Cambrian explosion
- Opabinia
- Wiwaxia
Footnotes
- 1 2 Daley, A. C.; Peel, J. S. (2010). "A Possible Anomalocaridid from the Cambrian Sirius Passet Lagerstätte, North Greenland". Journal of Paleontology. 84 (2): 352–355. doi:10.1666/09-136R1.1.
- 1 2 3 4 5 6 7 Conway Morris, S. (1998). The crucible of creation: the Burgess Shale and the rise of animals. Oxford [Oxfordshire]: Oxford University Press. pp. 56–9. ISBN 0-19-850256-7.
- 1 2 Whittington, H.B.; Briggs, D.E.G. (1985). "The largest Cambrian animal, Anomalocaris, Burgess Shale, British Columbia". Philosophical Transactions of the Royal Society B. 309 (1141): 569–609. Bibcode:1985RSPTB.309..569W. doi:10.1098/rstb.1985.0096.
- 1 2 3 Usami, Yoshiyuki (2006). "Theoretical study on the body form and swimming pattern of Anomalocaris based on hydrodynamic simulation". Journal of Theoretical Biology. 238 (1): 11–7. doi:10.1016/j.jtbi.2005.05.008. PMID 16002096.
- 1 2 3 Whittington, H.B.; Briggs, D.E.G. (1985). "The Largest Cambrian Animal, Anomalocaris, Burgess Shale, British Columbia". Philosophical Transactions of the Royal Society B (free full text). 309 (1141): 569–609. Bibcode:1985RSPTB.309..569W. doi:10.1098/rstb.1985.0096.
- 1 2 Briggs, Derek E. G. (1994). "Giant Predators from the Cambrian of China". Science. 264 (5163): 1283–4. Bibcode:1994Sci...264.1283B. doi:10.1126/science.264.5163.1283. PMID 17780843.
- ↑ John R. Paterson; Diego C. García-Bellido; Michael S. Y. Lee; Glenn A. Brock; James B. Jago; Gregory D. Edgecombe (2011). "Acute vision in the giant Cambrian predator Anomalocaris and the origin of compound eyes". Nature. 480 (7376): 237–240. Bibcode:2011Natur.480..237P. doi:10.1038/nature10689.
- ↑ "Ancient super-predator eyes found in Australia". Seven News. December 8, 2011. Retrieved December 11, 2011.
- 1 2 3 4 5 6 7 8 Gould, Stephen Jay (1989). Wonderful life: the Burgess Shale and the nature of history. New York: W.W. Norton. pp. 194–206. ISBN 0-393-02705-8.
- ↑ "The Anomalocaris homepage". Archived from the original on 4 April 2008. Retrieved 2008-03-20.
- ↑ Daley, A. and Bergström, J. (2012). "The oral cone of Anomalocaris is not a classic 'peytoia'." Naturwissenschaften, doi:10.1007/s00114-012-0910-8
- ↑ Ancient discovery puts world's scientific eyes on Kangaroo Island ABC News (Australia) 30 June 2011
- ↑ Salleh, Anna (December 8, 2011). "Cambrian predator had killer eyes". ABC Science. Retrieved 15 February 2012.
- ↑ Fossilised eyes of ancient super-predator found Australian Geographic December 9, 2011
- ↑ Briggs, D. E. G.; Mount, J. D. (1982). "The Occurrence of the Giant Arthropod Anomalocaris in the Lower Cambrian of Southern California, and the Overall Distribution of the Genus". Journal of Paleontology. 56 (5): 1112–8. JSTOR 1304568.
- 1 2 3 4 5 6 Nedin, C. (1999). "Anomalocaris predation on nonmineralized and mineralized trilobites". Geology. 27 (11): 987–990. Bibcode:1999Geo....27..987N. doi:10.1130/0091-7613(1999)027<0987:APONAM>2.3.CO;2.
- ↑ Briggs, D. E. G.; Robison, R. A. (1984). "Exceptionally preserved nontrilobite arthropods and Anomalocaris from the Middle Cambrian of Utah". University of Kansas Paleontological Contributions (111). ISSN 0075-5052.
- 1 2 Hagadorn, James W. (August 2009). "Taking a Bite out of Anomalocaris" (PDF). In Smith, Martin R.; O'Brien, Lorna J.; Caron, Jean-Bernard. Abstract Volume. International Conference on the Cambrian Explosion (Walcott 2009). Toronto, Ontario, Canada: The Burgess Shale Consortium (published 31 July 2009). ISBN 978-0-9812885-1-2.
- 1 2 Daley, A. C.; Paterson, J. R.; Edgecombe, G. D.; García-Bellido, D. C.; Jago, J. B. (2013). Donoghue, Philip, ed. "New anatomical information on Anomalocaris from the Cambrian Emu Bay Shale of South Australia and a reassessment of its inferred predatory habits". Palaeontology: n/a. doi:10.1111/pala.12029.
- ↑ Daley, A. C.; Bergström, J. (2012). "The oral cone of Anomalocaris is not a classic peytoia". Naturwissenschaften. 99 (6): 501–504. Bibcode:2012NW.....99..501D. doi:10.1007/s00114-012-0910-8. PMID 22476406.
- ↑ Vinther, J.; Stein, M.; Longrich, N. R.; Harper, D. A. T. (2014). "A suspension-feeding anomalocarid from the Early Cambrian". Nature. 507: 496–499. doi:10.1038/nature13010.
- ↑ Van Roy, P.; Daley, A.C.; Briggs, D.E.G. (2015). "Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps". Nature. 522: 77–80. doi:10.1038/nature14256.
- ↑ Witze, Alexandra, "Fossil fangs not so fierce", Science News, Vol.178 #11 (p. 13). Online edition retrieved 2010.11.11
External links
| Wikimedia Commons has media related to Anomalocaris. |
- "Anomalocaris canadensis". Burgess Shale Fossil Gallery. Virtual Museum of Canada. 2011.
- Anomalocaris 'homepage' with swimming animation
- Burgess Shale: Anomalocaris canadensis (proto-arthropod), Smithsonian.